New Representative of the Family Hydroscaphidae from Tunguska Basin, the Boundary of the Permian and Triassic (Coleoptera, Myxophaga) with Review on Myxophagan Fossil R ecords
Palaeoscapha tunguskaensis gen. et sp. nov., the oldest representative of the family Hydroscaphidae, is described from the Tunguska Basin in Siberia, dated from the boundary of the Permian and Triassic. It is assigned to the subfamily Triamyxinae, stat. nov. New diagnoses of the Hydroscaphidae and Triamyxinae are proposed. The new subfamily Leehermaniinae subfam. nov. (type genus Leehermania) is erected. The fossil record of the suborder Myxophaga is reviewed and revised with some taxonomic notes and synonymy of some generic and species names in the family Lepiceridae, known from Burmese amber. Different aspects of the evolutionary history of this suborder are discussed. A synonymy of Haplochelus and Lepiceratus is proposed.
## I. INTRODUCTION
like other myxophagan families, the Hydroscaphidae is a relatively small group in the Recent fauna, and it seems to be yet poorly known in fossils. The extant hydroscaphids were reviewed thanks to the efforts of many colleagues [4, 7, 9, 10, 23, 24, 25, 27, 28, 29, 35, 36, 37, 38, 39, 42, 45, 46, 47, etc.]. It currently comprises four genera and nearly 40 species [47], while the modern myxophagans include about 120 species in four families with apparently ancient origins. Recently one hydroscaphid specimen was found in materials in sediments from Tunguska Basin, Krasnoyarsky Kray (tentatively dated around the boundary between the Permian and Triassic). The suborder Myxophaga sensu stricto has only a few fossil urn:lsid:zoobank.org:pub:57B4F997-0BA9-45E0-8B8E-264FDEC4 C05E
records [3, 4, 8, 9, 12, 13, 15, 16, 18, 19, 26, 34], which need to be reviewed and discussed to clarify the composition of this group in the past. The paper is aimed to describe the oldest fossil myxophagan representative (Paleoscapha tunguskaensis gen. et sp. nov.), summarize the available data on the current fossil record of the myxophagans, and provide them with some essential comments, including the proposal of a new subfamily Leehermaniinae subfam. nov. (type genus Leehermania Chatzimanolis, Grimaldi, Engel et Fraser, 2012) [3], and a subfamily state for Triamyxidae Qvarnström, Fikáček, Wernström, Huld, Beutel, Arriaga-Varela, Ahlberg et Niedzwiedzki, 2021 (Triamyxinae stat. nov.) [26].
The suborder Myxophaga sensu stricto was proposed by Crowson [4] only for the groups from the Recent fauna with some characters similar to those in the suborder Archostemata (mostly apparently plesiomorphic), in contrast to others which are shared by both myxophagans and polyphagans and seem to be more derivative but probably homoplastic. The recent comprehensive studies of fossil beetles revealed close relations between the recent myxophagan families and extinct archostematan groups of the infraorder Schizosphoriformia Kirejtshuk, 1991 \[14\](=Schizosphorimorpha, Ponomarenko, 2001 [30]) [32]. In some recent publications both these groups are sometimes regarded together in the suborder Myxophaga sensu lato (e.g.,[1, 47]; etc.). The authors of this paper consider that such a joining remains still not reasonable and prefer to interpret them in the traditional way, viz. separating an archostematan infraorder Schizosphoriformia from a suborder Myxophaga sensu stricto).
## II. MATERIAL AND METHODS
The specimen examined was buried in volcanic rock and partially replaced by silica. As a result, the beetle remains maintains a three-dimensional appearance. This circumstance explains its comparatively good preservation. This fossil remains is one of two insects, beetle and leaf-hopper [44] found in
Untuun River outcrop. There is no findings of similar beetles among the oryctocenoses of other volcanogenic localities of the Siberian (including Tunguska) traps. The many kilometer thickness of traps could have been accumulated over a short time and consist of a sequence of explosive thermal tuffs and a sequence of erupted basaltic lavas. All crustaceans, insects, fishes, and tetrapods from these localities belong to the first sequence of the traps. The thermal effect of lavas on coals and carbonates of the Tunguska syncline probably led to the accumulation of atmospheric methane and carbon oxides, which led to the thermal maximum in the middle of the Induan time [50]. If these assumptions are correct, the temperature maximum has fallen at the end of the Induan. Therefore, the Permian-Triassic boundary on the Tunguska should be placed on the outpouring of basalts. In this case, all locations of fossil organisms on the Tunguska traps should be considered as early Triassic and Induan, as was usually treated before with reference to the character of their fauna and flora, and not Permian as suggested by Sadovnikov [40, 41].
The specimen under description was examined by A.G. Ponomarenko and A.A. Prokin using a Leica M16 light stereomicroscope with a Leica DFC420 digital camera and an attached Tescan Vega XMU scanning electron microscope. The line drawing was prepared using Adobe Photoshop CS3. Some additional fossil and recent specimens from the collections of the Zoological Institute of the Russian Academy of Sciences (St. Petersburg), Borissiak Paleontological Institute of the Russian Academy of Sciences (Moscow), and Muséum national d'Histoire naturelle (Paris) were studied for comparison of the specimen under description. Photographs of different focal depths were combined using Helicon Focus 5.1.28. Line drawing was prepared with the Autodesk AutoCAD 2015 software.
## III. SYSTEMATIC PALEONTOLOGY
Order Coleoptera Linnaeus, 1758
Suborder Myxophaga Crowson, 1955 [4]
Family Hydroscaphidae Le Conte, 1874 [22]
Remarks: This family was under the attention of many researches (see above); however, the current review of available myxophagan fossils forces us to recompile an emended diagnosis for this family.
Diagnosis (adults): Body small and fusiform, more or less gently narrowing to abdominal apex; usually dark and pubescent. Head large, wide and short, prognathous to hypognathous; eyes usually large, not protruding; antennae 5-11-segmented, usually clubbed; labrum separated from frons (clypeus); ultimate maxillary palpomere small, subulate; ultimate labial palpomere small and narrowing apically; mentum very small to relatively large. Prothorax rather short to moderately long; propleura reaching anterior edge of prothorax; prosternum relatively short and with short and pointed process; procoxal cavities transversely oval to transverse; procoxae with exposed trochantin. Mesothorax short or moderately long, with depressions for receipt base of prolegs; mesocoxal cavities widely to narrowly separated, externally opened; mesocoxae oval. Metathorax usually moderately long; metepisterna moderately widened anteriorly and well exposed; metaventrite without trace of sutures or with expressed premetacoxal sutures; metacoxae widely separated to sub (contiguous), usually short and with more or less raised plates or without them. Abdomen usually with six ventrites either rarely with five (probably in females) or seven ventrites, ventrite 1 longest (when all following segments of similar structure); 1-2 terminal segments subconical and very long. Elytra shortened and transversely truncate at apex or slightly shortened and subacute at apices; hind wing without oblongum, with reduced anal field and fringed along edge. Legs moderately long; trochanters elongate; femora of usual shape; tibiae relatively narrow to moderately wide or rarely strongly dilate; three simple tarsomeres.
Subfamily Triamyxinae Qvamström, Fikáček, Wernström, Huld, Beutel, Arriaga-Varela, Ahlberg et Niedźwiedzki, 2021 [26], stat nov.
Type genus. Triamyxa Qvarnström, Fikáček, Wernström, Huld, Beutel, Arriaga-Varela, Ahlberg et Niedźwiedzki, 2021 [26].
Remarks: The taxon 'Triamixidae' was proposed as a separate family among the suborder Myxophaga [26], based on characters from the list compiled by Lawrence et al. [20] and thanks to computer software for producing cladograms interpreted as follows: 'Bayesian analyses of Coleoptera and Myxophaga datasets with constrained molecular topology suggest a sister group relationship with the entire modern Myxophaga. Maximum parsimony analyses place it either in an unresolved basal polytomy (analyses with constrained molecular topology...) or as sister to Hydroscaphidae (unconstrained analyses). The character matrix adopted by Qvarnström et al. [26] for analyses and determination of the placement of the studied type specimens of the fossil species contain 77 characters extracted from the set of 359 characters elaborated and defined for modern species [21]. Qvarnström et al. [26] could not obtain adequate definitions of characters to compare fossil representatives with extant myxophagans. Besides, the latter researchers did not provide their description with a proper family diagnosis, and they published only some comparison ('differentiated diagnosis') after the formalization of character definition by Lawrence et al. [21] and 'table S1' with 'family' diagnosis and comparison with other groups of the suborder Myxophaga sensu stricto. The new comparison of myxophagan fossils allows us to join two fossil genera (Triamyxa and Palaeoscapha gen. nov.). The newly defined diagnostic group characters here revised gives a possibility to generate a new emended diagnosis for this suprageneric taxon with a new rank (subfamily).
Because many characters of this subfamily are poorly visible in the fossil compressions examined, the diagnostic group features of this taxon should be regarded as preliminary. They require further revision after future studies of new additional specimens. The large mentum is previously included in the triamyxid diagnosis, as it was observed in both Triamyxa and Palaeoscapha gen. nov. The precoxal part of prosternum is rather long in Triamyxa, but moderately long in Palaeoscapha gen. nov. Triamyxa has probably nine antennomeres with a three-segmented club, while the antennae in the holotype of Palaeoscapha tunguskaensis sp. nov. are missing. Therefore, the antennal characters in the subfamily diagnosis proposed below should be considered as putative for the new genus. The holotype of Palaeoscapha tunguskaensis sp. nov. has six exposed abdominal ventrites, while the specimens of the type series of Triamyxa coprolithica Qvarnstrom, Fikáček, Wernström, Huld, Beutel, Arriaga-Varela, Ahlberg et Niedźwiedzki, 2021 [26] have five or six exposed ventrites (\[26\]: 'females with 5 ventrites and males with 6 ventrites'). The reconstruction of the prothoracic structure made by Qvarnström et al. (2021) [26] needs some essential corrections. It was performed by forming a prothorax from separate pronotum and prosternum and then supposing that intervals between them could be interpreted as a space for propleura (Qvarnström et al., 2021 \[26\]: Fig. 3, R-T). However, it is visible that the left mesal outline of the pronotum in Fig. 3T is different from the right mesal outline of the same pronotum. Furthermore, if all three pronota (Fig. 3 R-T) are compared, it can be supposed that a narrow interval between the anterior mesal angle of the pronotum and the anterior lateral angle of the prosternum is quite acceptable. If so, the diagnostic character of Hydroscaphidae concerning the propleura reaching the anterior edge of the prothorax can certainly be defined as in the below diagnosis.
Diagnosis: Head apparently prognathous, with eyes located at base moderately arcuately protruding and rather short temples not projecting laterally. Antenna clubbed with apparently nine antennomeres. Mentum very large. Mesothorax comparable in length with prothorax. Metepisterna moderately and almost rectilinearly anteriorly widened, their mesal anterior angle very narrowly separated from outer edge of mesocoaxal cavity. Metacoxae very narrowly separated to conjoining and without raised coxal plates. Elytra full to slightly shortened and narrowly transversely truncate or subacute at apex; epipleura extending behind level of metacoxae. Abdomen with 5-6 ventrites.
Composition: The type genus (Triamyxa) and Palaeoscapha gen. nov.
Genus Palaeoscapha Kirejtshuk, Prokin et Ponomarenko, gen. nov.
http://zoobank.org/urn:lsid:zoobank.org:act:05BBDE9D-F07C-4659-BF0F-B4F83BDBF5DE
Type species: Palaeoscapha tunguskaensis sp. nov.
Etymology: The name of this new genus is formed from the Greek $\pi \breve{\alpha} \lambda \alpha \iota \varsigma =$ palaeo' (ancient, older) plus generic root 'scapha' (Scapha, Hydroscapha, Scaphander, Scaphydra). Gender feminine.
Remarks: The new genus is represented only by one species and, therefore, the diagnoses of both somehow coincide and can be used for both species and genus in accordance of the traditional principle 'descriptio generica specifica'.
Diagnosis: Body somewhat elongate, rather convex ventrally. Head subtriangular, apparently prognathous and truncate at anterior edge of frons (clypeus); gular sutures arcuately divergent; eyes located at base and medium-sized, moderately protruding, from below looking like equally biconvex lens; mandibles apparently slightly raised and apparently arcuate along outer edge; mentum large, widest at base and arcuately narrowing anteriorly; antennal grooves distinct, arcuate and reaching level of posterior edge of mentum; temples arcuately narrowing posteriorly. Prothorax transverse, widest at widely rounded posterior angles, gently narrowing to apparently slightly projecting anterior angles; prosternum very short and subtriangular, prosternal process very narrow, short and nearly sharply acuminate at apex, reaching midlength of procoxae; propleura reaching anterior edge of prothorax, apparently with mesal process extending along anterior edge of procoxae; procoxal cavity large, transverse and seemingly open posteriorly. Procoxae narrowly separated rather than contiguous, subacutely angular laterally and with largely exposed trochantin. Mesepisternum and mesepimeron comparable in size; mesoventrite subequal in length with prothorax; mesocoaxal cavity open externally. Mesocoxae large, suboval, comparatively narrowly separated. Metathorax somewhat longer than each of pro- or mesothoraces; metaventrite transverse and with paramedian nearly rectilinear lines from inner edge of mesocoxae to outer edge of metacoxae; premetacoxal sutures slightly expressed and subparallel to anterior edge of metacoxal cavities. Metacoxae relatively short, (sub)contiguous and rectilinearly located along entire posterior edge of metaventrite. Elytra covering abdomen, about 1.5 times as long as wide together and subangular at apex;
epipleura reaching at least level of ventrite 1. Abdomen with six ventrites and comparatively short, only slightly longer than wide; ventrite 1 longest and about twice as long as each of ventrites 2-6; hypopygidium (ventrite 6) short, transverse and widely rounded at apex. Right anterior leg with elongate trochanter, femur of very usual shape and moderately wide; protibia dilated along inner side and relatively wide.
Note: Posterior edges of abdominal ventrites looking like even (without notches).
Comparison: The new genus differs from all hydroscaphids in the dilated protibiae and relatively short abdomen with all ventrites strongly transverse; and also from Triamyxa in the larger mentum, smaller eyes, distinct and curved antennal grooves, transverse procoxae, widely rounded posterior angles of the pronotum and subacute elytral apices.
Palaeoscapha tunguskaensis Kirejtshuk, Prokin et Ponomarenko, sp. nov. Figs 1-3 http://zoobank.org/urn:lsid:zoobank.org:act: 37EAED97-A725-4CCA-8625-AE1F2303B33F
Type material: Holotype, sex unknown; specimen represented by three-dimensional remains mostly visible from underside, PIN no. 2757/1, demonstrating the most part of its body, although the right part of the pro- and pterothoraces, abdominal apex, and appendages are missing (except for the observable right anterior trochanter, femur and tibia). The pictures of the holotype were published by Kirejtshuk & Ponomarenko[20] and Ponomarenko & Prokin[34].
Type locality and horizon: Russia, Krasnoyarsky Kray, Evenkiysky National Okrug, Tunguska Basin, Untuun River; intertrappean deposits; probably early Triassic or terminal Permian, Induan or?Tatarian.
Etymology: The epithet of the new species is formed from the name of its type locality (Tunguska).
Description (holotype): Body length 1.5, body width 0.6 mm. Integument mostly smoothed, moderately finely and sparsely punctured (puncture diameter about as great as that of eye facets); prosternum transversely striate, propleura rugose, ventrites partly shagreened. Head subtriangular, apparently prognathous; distance between gular sutures subequal with maximum width of mentum and smaller than that between eyes; mentum slightly transverse, trapezoid, subtruncate at base and slightly arcuately narrowing anteriorly. Precoxal part of prostenum noticeably shorter than mesal length of procoxae. Mesocoxae about twice as widely separately as procoxae. Metaventrite slightly transverse; metepisterna subtriangular; premetacoxal sutures slightly expressed closely subparallel to anterior edge of metacoxal cavities. Elytral integument seemingly smoothed.
## IV. DISCUSSION
The subfamily Hydroscaphinae of the family Hydroscaphidae of the superfamily Sphaeriusoidea was known only after the one compression fossil from the Cretaceous of the Yxian Formation (Huangbanjigou, Liaoning, China) [2]. However, most outlines of the sclerites of this fossil are rather unclear. Therefore, the suborder and family attributions of this taxon should be regarded as rather probable. The distinctness of separated meso- and metacoxae, and also three-segmented tarsi, which are used by the co-authors of this publication as the most essential arguments, can be only traced with some probability. This fossil has not enough characters to be compared with modern genera and to justify its attribution to Hydroscapha rather than to any other genus in Hydroscaphidae. Thus, the species name 'jeholensis' should be attributed rather to a genus incertus or cf. Hydroscapha than strictly to the latter genus. Nevertheless, the assignment of this species to the subfamily Hydroscaphinae sensu stricto is clear enough after the key to the hydroscaphid subfamilies proposed below.
The second hydroscaphid subfamily Triamyinae stat. nov. is extinct and known only after two findings from the Permian-Triassic boundary (Palaeoscapha gen. nov.) and late Triassic (Triamyxa). It is characterized in detail in the above diagnosis and remarks, and can be easily identified after the below key to hydroscaphid subfamilies.
The late Triassic Leehermania prorova Chatzimanolis, Grimaldi, Engel et Fraser, 2012 [3] from Cascade (near Martinsville, Virginia, USA; Cow Branch Formation, late Carnian - early Norian), originally described as the oldest Staphylinidae [3], was recently re-examined and re-interpreted as a probable 'ancestral' Hydroscaphidae [8]. The type species of this genus seems to have enough characters to put the genus Leehermania near the subfamily Triamyxinae stat. nov., but this genus, in contrast to Triamyxa and Palaeoscapha gen. nov. (see the above remarks to the subfamily Triamyxinae stat. nov.), has a markedly hypognathous head, pronotum widest at the midlength, relatively short meso- and metathoraces, very short elytra with truncate apices and seven abdominal ventrites with comparatively shorter ventrite 1. These mentioned differences make it possible to propose for this genus a new subfamily. Both descriptors [3] and re-examinators [8] of Leehermania indicated the presence of about six ventrites (derivatives of sternites III-VIII), while the pictures of lateral view in both publications clearly show seven segments on the underside of the abdomen, although Chatzimanolis et al. [3] pointed out that the 'lateral tergal sclerites of IX becoming tapered to narrow apex'. It is a rather important point that the specimens of Leehermania prorova have two basal ventrites vs. only one as indicated by Fikáček et al. [8] in the drawing-reconstruction of the Fig. 2H of their publication. This misinterpretation can be evident even comparing this reconstruction with the picture of the specimen used for this drawing-reconstruction (Fig. 2G). It is also clearly visible in Chatzimanolis at al. (\[3\]: Fig. 8). Besides, the abdominal ventrite 1 of Leehermania prorova has its length only slightly more significant than the length of the ventrites 2-6 (the proportion of the ventrite 1 is significantly greater in all other known hydroscaphids). The same also concerns the hypognathous head of Leehermania prorova, although some modern hydroscaphids could have their heads in hypognathous positions. The abovementioned arguments show a considerable distinctness of the considered species, which is treated as enough ground to propose a new extinct subfamily Leehermaniinae subfam. nov.
The second sphaeriusoid family is Sphaeriusidae Erichson, 1845 [6], also known in the fossil record as a single species from Burmese amber (Burmasporum rossi) described by Kirejtshuk [15] with 'generalized' structural features compared to the modern representatives of the family (members of the genus Sphaerius Erichson, 1845 [6]). It seems to have many possible plesiomorphic differences from the modern species, namely: the comparatively larger, longer and more projecting head with longer anterior part of frons and longer mouthparts, and distinctly elongate eyes; elongate scape and antennomere 2, small antennomere 3, loose antennal club with four antennomeres; movable and larger pronotum with shorter lateral parts and widely rounded anterior and posterior angles and comparatively long legs with simple and subparallel-sided femora and tibiae.
The super family Lepiceroidea is represented by the single family Lepiceridae in the Recent fauna. Some fossil representatives of this family are known from the Cretaceous Burmese amber. At first, this family had only three fossil specimens from the same source (Burmese amber), and all these specimens are rather poorly preserved for studies, so that some interpretations and conclusions appear rather ambiguous and partly inadequate because of the not quite clearly visible structures examined. On the other hand, the studies of these specimens were rather essential in understanding the myxophagan phylogeny. Therefore, the conclusions obtained after studies of the first two specimens by Kirejtshuk and Poinar [18, 19], and the conclusions obtained after comparison of the third specimen with previous information by Jałoszyński et al. [12] are different. The latter co-authors tried to find a compromise based on the available prior knowledge using the particular matrix of the characters for a cladogram or on an assumption that the facts of fossil representatives are essentially wrong [12]. As a result, the co-authors of the last publication regarded that all Mesozoic species belong to the same genus, which is also present in the modern fauna (Lepicerus). They treated the description of Haplochelus as obscure and unworthy. They decided that the diagnostic characters of both fossil genera Haplochelus Kirejtshuk et Poinar, 2006 [18] and Lepichelus Kirejtshuk et Poinar in Kirejtshuk, 2017 \[16\](=Lepiceroides Kirejtshuk et Poinar, 2013 [19], non Schedl, 1957 [43]), and also modern Lepicerus could be treated as only specific. However, most characters of Lepichelus pretiosus (Kirejtshuk et Poinar 2013) [19], comb. nov. and Lepichelus mumia (Jaloszyński et Yamamoto in Jaloszyński et al., 2017) [12], comb. nov. are correspondent to each other. Differences between them could be depended upon preservation condition of examined specimens. The situation became more confusing because these co-authors some times declared that 'Lepicerus' mumia has four antennomeres, as, in their opinion, should be in the modern members of Lepicerus, but the drawing in their paper (Fig. 2D) clearly shows five antennomeres as should be in Lepichelus (indeed according to Fikáček et al. [8] modern members of Lepicerus have four antennomeres in one species and five in others). Thus, the contradictions between the opinions of researchers can be proved or disproved only after the examination of additional specimens and further examination of the holotypes. The description of the poorly preserved material of Lepichelus mumia comb. nov. does not allow to clearly discriminate it from L. pretiosus comb. nov. [16]. The 'curved protarsomere 1' mentioned by Fikáček et al. [8] for Lepichelus mumia comb. nov. could be indeed a result of an optic aberration. These codescribers also indicated the following characters of Lepichelus mumia comb. nov. diagnostic from 'L. pretiosus, in the epipleural concavity located behind the middle of the elytra (at the middle in L. pretiosus), and in the prosternal process narrowing, and not broadening posteriorly'. Perhaps, the problem of the conspecificity of specimens and synonymy of these species names has to be finally solved after a further re-examination of both type specimens and additional specimens. The extinct Cretaceous species differ from extant ones in the structure of elytral sides and less widely separated metacoxae (almost about comparably separated with mesocoxae). Anyway, it seems to be more correct to consider that the palaeoendemic genus Lepichelus comprising two species from Burmese amber are enough separated from the taxon with modern representatives, and it is not reasonable to join the modern and Cretaceous species in one generic taxon or tentatively consider them as two subgenera of the same genus.
The recent publication by Jałoszyński et al. [13], unfortunately, also contains some new wrong interpretations, but the good illustrations make it possible to solve some central problems in the contradictions and interpretations. In the last paper, the co-authors described the new genus Lepiceratus Jałoszyński, Luo, Yamamoto et Beutel, 2020 [13], which indeed completely fits with Haplochelus even in the number of antennomeres, the structure of elytra, comparatively narrowly separated all pairs of coxae of the type specimen of the genus type species (L. ankylosaurus Jałoszyński, Luo, Yamamoto et Beutel, 2020 [13]). The type species of both genera (Haplochelus and Lepiceratus) have some differences in the general body outline, different shapes of head and pronotum, eye sizes, presence of the visible discrimen only in $H$. georyssoides vs. absence in $L$. ankylosaurus, and some other features, which are better to consider as species differences than generic ones. Even the elytra of the latter species (L. ankylosaurus) are distinctly separated only in the artist's reconstruction of $L$. ankylosaurus (\[13\]: Fig. 1C) but not so distinctly in the photograph (Fig. 1A) and tomography reconstruction (Fig. 2A and 3A). It is thought that the sutural elytral edges can be relatively sharply elevated and firmly closed together to make an impression of the united fused ridge (as in the holotype of Haplochelus georyssoides). A median depression of the mentum with the sharp outline of this holotype could make an illusion of gular sutures [18], and also a more or less weak median depression is quite characteristic of other lepicerids. Thus, it is reasonable to consider the name Lepiceratus as a junior synonym of Haplochelus.
Taking into consideration the small body size of all known extant and fossil myxophagans and a small probability of their coming into deposits (tanatocenoses), it can be supposed that the available fossil record of this group is rather scarce among coleopterans in general. Nevertheless, the suborder Myxophaga is rather ancient and originated not later in the Palaeozoic (as suborders Archostemata and Adephaga). D.E. Shcherbakov found one unique tiny disc-like beetle larva in the extensive collection from the Middle Permian ( $\sim$ 265 Ma) of Kargala near Orenburg, European Russia (Borissiak Paleontological Institute, Russian Academy of Sciences), whose print demonstrates a considerable similarity to larvae of some modern members of Torridincolidae Steffan, 1964 \[49\](superfamily Sphaeriusoidea). The current data on this suborder is not sufficient enough for grounded phylogenetic conclusions. The rather ancient origin of Myxophaga and the scarcity of its fossil record make it impossible to use the current methods to reconstruct its phylogeny. It is thought that the groups of four myxophagan families in the Recent fauna represent only rather separate splinters of the significant diversity of this suborder in the past, although hydroscaphids (and probably something like torridincolid-like larvae) could be present in the Palaeozoic faunas. Other two myxophagan families (Sphaeriusidae and Lepiceridae) and also true torridincolids with more specialized structural appearance could have the later (Mesozoic) origin (as traced in the current fossil record of the suborder). Nevertheless, many phylogenetic hypotheses after formal analyses of structural and molecular comparisons give a basal placement for the Hydroscaphidae ([8, 26]; etc.), i.e., showing some correspondence with the current fossil record. It is necessary to take into consideration that the result of structural analysis is somehow connected with the more generalized appearance of hydroscaphids (a greater proportion of plesiomorphies in general). At the moment, strict phylogenetic reconstructions are somewhat premature. However, it can be supposed that the phylogeny of this suborder could have a complexity analogous to that of cupedids from the suborder Archostemata. A grounded phylogenetic model will be more probable after getting more data on fossils of various ages and applying the principle of multiple parallelisms [17].
The very conspicuous peculiarity of this suborder was pointed out by Crowson [5] that the myxophagan families have structural and partly bionomically analogous groups among various small polyphagans. In particular, adults of modern Hydroscaphidae, according to the opinion of R.A. Crowson, have many similarities with 'staphylinoids', 'hydrophiloids', and basalmost eucinetids [5]. The lepicerids, in general appearance, resemble elmids [5] or georissids [18]. The species of Sphaeriusidae somehow are reminiscent of polyphagan clambids (Scirtoidea) and cybocephalines (Nitidulidae, Cucujoidea) (see notes on 'Calyptomeridae' in Crowson, 1967 [5]). Thus, in the phylogenetic reconstruction of myxophagans, it would be desirable to take into consideration many aspects of structural convergence, including adaptive and morphogenetic processes.
It can be supposed some kind of association between Palaeozoic and Mesozoic myxophagans and algae, which is somewhat similar to that between the modern representatives of this suborder and their food resource. As another Permian suborder Archostemata with little structural transformations reached nowadays mostly because of the maintenance of ancient lifestyle and habits, modern myxophagans could also have a considerable conservatism in their habitats and diet from the Palaeozoic together with their general appearance, keeping small body-size and simplification of some structures. Modern myxophagans with known bionomy, including hydrosaphids, usually inhabit aquatic to moist environments associated with algal growth [47]. Probably a certain similarity in throphic connections should also be expected in the case of the fossil Palaeoscapha tunguskaensis gen. et sp. nov. As to Triamyxa coprolithica, described from coprolite of dinosaur form Silesaurus opolensis from the lacustrine Krasiejów deposits [26], it is thought that this beetle inhabited aquatic and moist environments associated with algal mats (most likely growing at shores of water bodies or floating). Floating mats or their aggregates are known from different Mesozoic localities and proposed as probable habitats for many aquatic insects, including beetles, as well as food resources for vertebrates such as dinosaurs [31, 33, 34].
## V. TAXONOMIC CONCLUSIONS
The available data of fossil records on Myxophaga, taking into consideration the above discussion, can be summarized as follows:
Superfamily Sphaeriusoidea Erichson, 1845 [6]
Family Hydroscaphidae LeConte, 1874 [22]
Subfamily Hydroscaphinae LeConte, 1874[22]
- (?) 'Hydroscapha' jeholensis Cai, Short et Huang, 2012 [2] - China, Liaoning Province (Huangbanjigou), Yixian Formation; Lower Cretaceous, Lower Aptian
- Subfamily Triamyinae Qvarnström, Fikáček, Wernström, Huld, Beutel, Arriaga-Varela, Ahlberg et Niedźwiedzki, 2021 [26], stat. nov.
- Triamyxa coprolithica Qvarnström, Fikáček, Wernström, Huld, Beutel, Arriaga-Varela, Ahlberg et Niedźwiedzki, 2021 [26] - Krasiejów clay pit (near Ozimek), Upper Silesia, Poland, Upper Triassic
- Palaeoscapha tunguskaensis gen. et sp. nov. - Siberia, Tunguska Basin, Untuun; probably early Triassic or terminal Permian, Induan or?Tatarian
Subfamily Leehermaniinae Kirejtshuk, Prokin et Ponomarenko, subfam. nov.
urn:lsid:zoobank.org:act:3B6B5BDD-1CD8-40AC-BFDB-461C76B23842
Type genus Leehermania prorova Chatzimanolis, Grimaldi, Engel et Fraser, 2012 [3]
Diagnosis. Head seemingly hypognathous with eyes located at base moderately arcuately protruding and very short temples. Antennae 11-segmented and clubbed. Mesothorax extremely short. Metathorax comparatively short. Metepisterna strongly and almost rectilinearly widened anteriorly, their mesal anterior angle very narrowly separated from outer edge of mesocoxal cavity. Metacoxae seemingly very narrowly separated to conjoining and apparently without raised coxal plates. Elytra somewhat shortened and widely transversely truncate at apex; epipleura seemingly not extending behind level of metacoxae. Abdomen with seven ventrites.
Leehermania prorova Chatzimanolis, Grimaldi, Engel et Fraser, 2012[3] - USA, Virginia, Cascade (near
Martinsville), Cow Branch Formation, Upper Triassic, Upper Carnian/Lower Norian mily Sphaeriusidae Erichson, 1845 [6]
- Burmasporum rossi Kirejtshuk 2009 [15] - Myanmar, Burmese amber; Myanmar, Kachin, Hukawng Valley; 'mid'-Cretaceous, Albian/Ceno-manian
Superfamily Lepiceroidea Hinton, 1936[11] Family Lepiceridae Hinton, 1936 [11]
- Lepichelus pretiosus (Kirejtshuk et Poinar, 2006), comb. nov. [19] [Lepiceroides] - Burmese amber; Myanmar, Kachin, Hukawng Valley; 'mid'Cretaceous, Albian/Cenomanian
- Probable synonym Lepichelus mumia (Jaloszynski and Yamamoto, 2017), comb. nov. [12] [Lepicerus] - Burmese amber; Myanmar, Kachin, Hukawng Valley; 'mid'-Cretaceous, Albian/Cenomanian
- Haplochelus georyssoides Kirejtshuk et Poinar, 2006 [18] - Burmese amber; Myanmar,Kachin, Hukawng Valley; 'mid'-Cretaceous, Albian/Cenomanian
- Haplochelus ankylosaurus (Jałoszyński, Luo, Yamamoto et Beutel, 2020), comb. nov. [13] \[Lepiceratus Jałoszyński, Luo, Yamamoto et Beutel, 2020, syn. nov. [13]\] - Burmese amber; Myanmar, Kachin, Hukawng Valley; 'mid'-Cretaceous, Albian/Cenomanian.
## VI. KEY TO HYDROSCAPHID SUBFAMILIES
1. Elytra long, covering completely or most part of abdomen and subacute at apices; abdomen with five or six ventrites; -metacoxae (sub) contiguous; mentum large; antennae with three-segmented club.Triamyxinae stat. nov.
- Elytra very short and truncate at apices, leaving 3-4 abdominal segments uncovered; abdomen with six or seven ventrites; - other characters different 2
2. Abdomen with six ventrites; antennae with five or nine antennomeres with ultimate sclub-shaped; metacoxae with coxal plate and clearly separated.. Hydroscaphinae sensu stricto
- Abdomen with seven ventrites; antennae with three-segmented club; metacoxae without coxal plate and narrowly separated or (sub) contiguous.......................................................... Leehermaniinae subfam. nov.
### ACKNOWLEDGEMENTS
The authors are grateful to R.A. Rakitov (Borissiak Paleontological Institute, Moscow, Russia) for help with SEM photos preparation and E.A. Prokina (Voronezh, Russia) for the help with pictures processing in Autodesk AutoCAD software, and particularly to D.E. Shcherbakov (Borissiak Paleontological Institute, Moscow, Russia) for various assistance, including seeking of specimens in the collection, picturing of fossil specimens, information on the larva from Kargala, valuable comments and help in providing the authors of this paper with literature. A. Nel (Mésem national d'Histoire naturelle, Paris) check the manuscript of this paper and made many very valuable comments and recommendations. The authors thank also anonymous reviewers and the editor for criticism of this manuscript during preparation and editing.
Funding
The studies of AGK were carried out under the framework of the Russian state research project no 1021051302540-6 and studies of AAP no 121051100109-1.
## APPENDIX A
After acceptance of this paper (November 23, 2022) two new papers on the fossil members of Sphaeriusidae appeared (December 7, 2022 and January 11, 2023):
The first paper has a description of the sphaeriusid genus Bezesporum Fikáček, Yamamoto, Matsumoto, Beutel et Maddison, 2022 (type species: Sphaerius minutus Liang et Jia, 2018 - modern, China, Jiangxi Province, Xiping County) including as two modern species as one fossil Bezesporum burmiticum



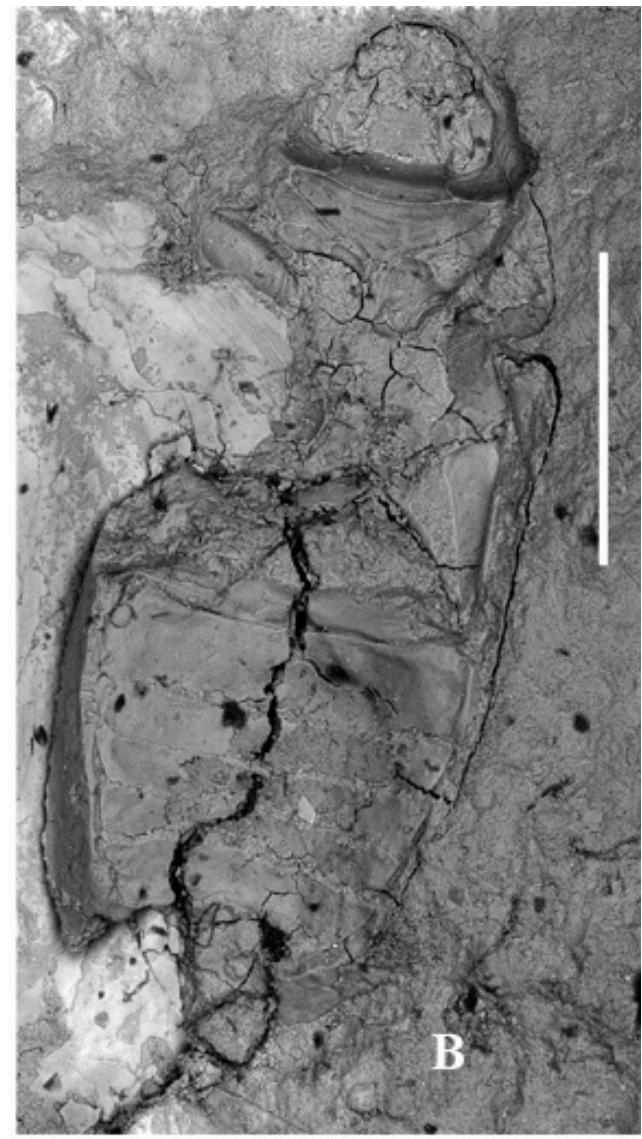
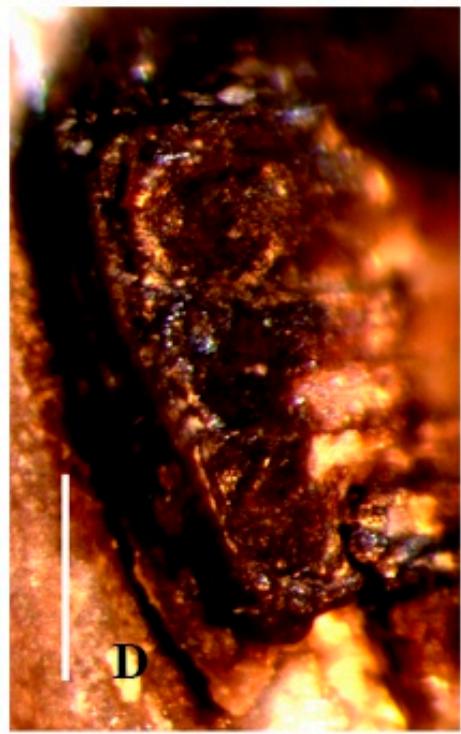 Figure 1: Palaeoscapha tunguskaensis gen. et sp. nov., holotype, photographs: A, B - habitus, C - head and thorax, D - abdomen. Scale bar 0.5 mm (A, B), 0.2 mm (C,D).

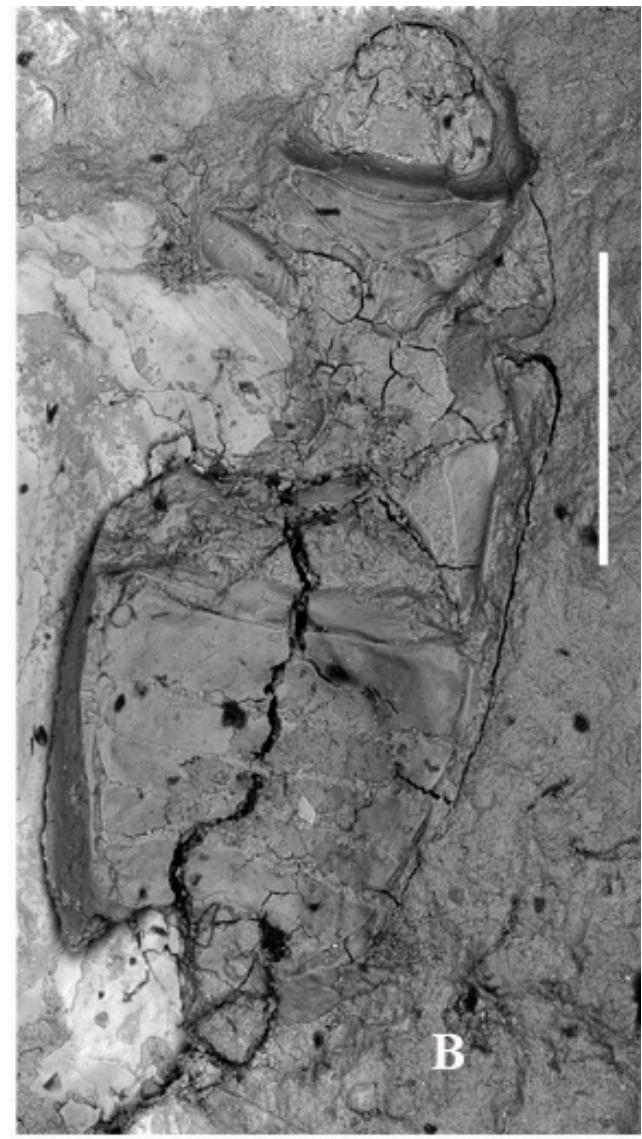

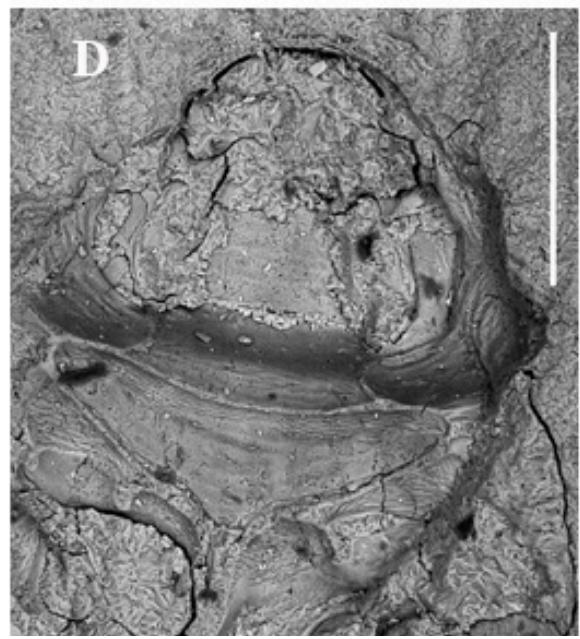 Figure 2: Palaeoscapha tunguskaensis gen. et sp. nov., holotype, SEM: A, B - habitus, C, D - head and prosternum. Scale bar 0.5 mm (B), 0.2 mm (D), not for scale (A,C).
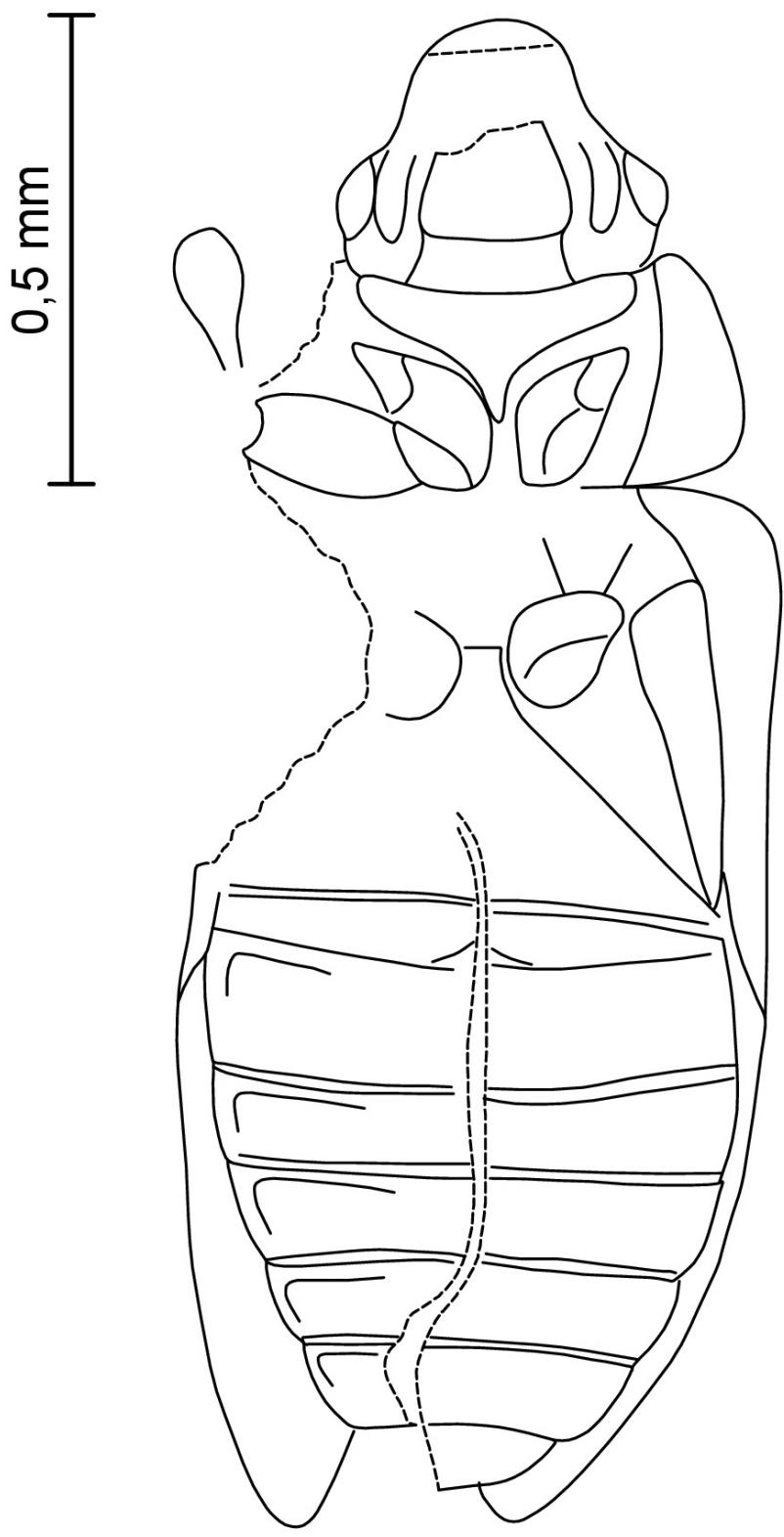 Figure 3: Palaeoscapha tunguskaensis gen. et sp. nov.: holotype, habitus, line drawing. Scale bar 0.5 mm.
Generating HTML Viewer...
References
51 Cites in Article
Patrice Bouchard,Yves Bousquet,Anthony Davies,Miguel Alonso-Zarazaga,John Lawrence,Christopher Lyal,Alfred Newton,Chris Reid,Michael Schmitt,Adam Slipinski,Andrew Smith (2011). Family-Group Names In Coleoptera (Insecta).
C Cai,A Short,D Huang (2012). The first skiff beetle (Coleoptera: Myxophaga: Hydroscaphidae) from Early Cretaceous Jehol biota.
Stylianos Chatzimanolis,David Grimaldi,Michael Engel,Nicholas Fraser (2012). <i>Leehermania prorova</i>, the Earliest Staphyliniform Beetle, from the Late Triassic of Virginia (Coleoptera: Staphylinidae).
R Crowson (1955). The natural classification of the families of Coleoptera.
R Crowson (1967). The natural classification of the families of Coleoptera.
W Erichson (1845). Unknown Title.
S Falamarzi,A Putz,M Heidari,H Nassersadeh (2010). Acta Entomologica Musei Nationalis Pragae.
Martin Fikáček,Rolf Beutel,Chenyang Cai,John Lawrence,Alfred Newton,Alexey Solodovnikov,Adam Ślipiński,Margaret Thayer,Shûhei Yamamoto (2019). Reliable placement of beetle fossils via phylogenetic analyses – Triassic <i>Leehermania</i> as a case study (Staphylinidae or Myxophaga?).
Martin Fikáček,Hana Šípková (2009). New Asian Hydroscapha, with comments on male-female association of co-occuring species (Coleoptera, Myxophaga, Hydroscaphidae).
W Hall,Andrew Short (2010). A new species of Hydroscapha LeConte from northwestern Venezuela(Coleoptera: Myxophaga: Hydroscaphidae).
Paweł Jałoszyński,Shûhei Yamamoto,Yui Takahashi (2017). Discovery of a new Mesozoic species of the ancient genus Lepicerus (Coleoptera: Myxophaga: Lepiceridae), with implications for the systematic placement of all previously described extinct ‘lepiceroids’.
Paweł Jałoszyński,Xiao-Zhu Luo,Jörg Hammel,Shûhei Yamamoto,Rolf Beutel (2020). The mid-Cretaceous †<i>Lepiceratus</i>Gen. nov. and the Evolution of the Relict Beetle Family Lepiceridae (Insecta: Coleoptera: Myxophaga).
A Kirejtshuk (1991). Evolution of mode of life as the basis for division of the beetles into groups of high taxonomic rank.
A Kirejtshuk (2009). A new genus and species of Sphaeriusidae (Coleoptera, Myxophaga) from Lower Cretaceous Burmese amber.
A Kirejtshuk (2017). Taxonomic notes on fossil beetles (Insecta: Coleoptera).
Alexander Kirejtshuk (2021). On subfamily structure of the Cupedidae (Coleoptera, Archostemata): data from paleontology and an approach to solving conflicting classifications.
A Kirejtshuk,G Poinar (2006). Haplochelidae, a new family of Cretaceous beetles (Coleoptera, Myxophaga) from Burmese amber.
A Kirejtshuk,G Poinar (2013). On the systematic position of the genera Lepiceroides gen.n. and Haplochelus, with notes on the taxonomy and phylogeny of the Myxophaga (Coleoptera).
Howard Hinton (1936). XLVI.—Lepiceridæ—<i>a new name for the</i>Cyathoceridæ. Lepicerinus—<i>a new name for the Scolytid genus</i>Lepicerus<i>Eichh. (Coleoptera)</i>.
A Kirejtshuk,A Ponomarenko (2008). Taxonomic notes on fossil beetles (Insecta: Coleoptera).
John Lawrence,Adam Ślipiński,Ainsley Seago,Margaret Thayer,Alfred Newton,Adriana Marvaldi (2011). Phylogeny of the Coleoptera Based on Morphological Characters of Adults and Larvae.
J Leconte (1874). Descriptions of New Coleoptera Chiefly from the Pacific Slope of North America.
I Löbl (1994). Les espèces asiatiques du genre Hydroscapha LeConte (Coleoptera, Hydroscaphidae).
Y Miwa (1935). Description of a new species of Hydroscaphidae from Formosa.
A Orchymont (1945). Notes sur le genre Hydroscapha LeConte (Coleoptera Polyphaga Staphyliniformia).
Martin Qvarnström,Martin Fikáček,Joel Vikberg Wernström,Sigrid Huld,Rolf Beutel,Emmanuel Arriaga-Varela,Per Ahlberg,Grzegorz Niedźwiedzki (2021). Exceptionally preserved beetles in a Triassic coprolite of putative dinosauriform origin.
Paul Peyerimhoff (1922). Nouveaux Coléoptères du Nord-Africain. Quarante et unième note : Hydroscaphidae.
Philip Perkins,Johannes Bergsten (2019). New Myxophagan water beetles from Madagascar (Coleoptera: Torridincolidae, Hydroscaphidae).
R Paulian (1949). Recherches sur les insectes d'importance biologique de Madagascar. VI. Deux familles de Coléoptères nouvelles pour la faune malgache.
A Ponomarenko (1973). On division of the order Coleoptera into suborders.
A Ponomarenko (1996). Evolution of continental aquatic ecosystems.
Julia Ponomarenko,Tatyana Merkulova,Galina Orlova,Oleg Fokin,Elena Gorshkov,Mikhail Ponomarenko (2002). Mining DNA sequences to predict sites which mutations cause genetic diseases.
A Rasnitsyn (1781). Class Insecta Linné, 1758. The Insects (=Scarabaeoda Laicharting, 1781).
A Ponomarenko,A Prokin (2015). Review of paleontological data on the evolution of aquatic beetles (Coleoptera).
L Pu (1948). A new species of Hydroscaphidae from China (Coleoptera).
H Reichardt (1971). Three new Hydroscaphidae from Brazil (Coleoptera, Myxophaga).
Hans Reichardt (1973). A critical study of the suborder Myxophaga, with a taxonomic revision of the Brazilian Torridincolidae and Hydroscaphidae (Coleoptera).
Hans Reichardt (1974). A critical study of the suborder Myxophaga, with a taxonomic revision of the Brazilian Torridincolidae and Hydroscaphidae (Coleoptera).
G Sadovnikov (2015). Paleoecological characterization of the Middle Siberian trappean plateau during the middle period of its formation (terminal Permian).
G Sadovnikov (2016). Evolution of the biome of the Middle Siberian Trappean Plateau.
M Satô (1972). A New Species of Stick Insect (Phasmatodea:Heteropterygidae:Obrimini) from Cebu Island, Philippines.
K Schedl (1952). Scolytoidea nouveaux du Congo Belge II: Mission R. Mayne -K. E. Schedl.
D Shcherbakov (2022). New Dysmorphoptilidae (Cicadomorpha) from the end-Permian and Middle Jurassic of Siberia: earliest evidence of acoustic communication in Hemiptera and the latest find of the family.
H Reichardt,H Hinton (1976). On the New World beetles of the family Hydroscaphidae. Papéis Avulsos de Zoologia 30, 1-24. communication in Hemiptera and the latest find of the family.
Andrew Short,Luis Joly,Mauricio García (2010). Discovery of the Skiff Beetle Genus<i>Yara</i>Reichardt and Hinton (Coleoptera: Myxophaga: Hydroscaphidae) in Venezuela, with Description of a New Species.
Andrew Short (2013). Aquatic Beetles of the Grensgebergte and Kasikasima Regions, Suriname (Insecta: Coleoptera).
Andrew Short,Luis Joly,Mauricio García,Alexander Wild,Devin Bloom,David Maddison (2015). Molecular phylogeny of the <scp>H</scp> ydroscaphidae ( <scp>C</scp> oleoptera: <scp>M</scp> yxophaga) with description of a remarkable new lineage from the <scp>G</scp> uiana <scp>S</scp> hield.
S Slipinski,R Leschen,J Lawrence (2011). Order Coleoptera Linnaeus, 1758. In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness.
A Steffan (1964). Torridincolidae, coleopterorum nova familia e regione aethiopica.
Yadong Sun,Michael Joachimski,Paul Wignall,Chunbo Yan,Yanlong Chen,Haishui Jiang,Lina Wang,Xulong Lai (2012). Lethally Hot Temperatures During the Early Triassic Greenhouse.
No ethics committee approval was required for this article type.
Data Availability
Not applicable for this article.
How to Cite This Article
Alexander G. Kirejtshuk. 2026. \u201cNew Representative of the Family Hydroscaphidae from Tunguska Basin, the Boundary of the Permian and Triassic (Coleoptera, Myxophaga) with Review on Myxophagan Fossil R ecords\u201d. Global Journal of Science Frontier Research - C: Biological Science GJSFR-C Volume 22 (GJSFR Volume 22 Issue C3).
Explore published articles in an immersive Augmented Reality environment. Our platform converts research papers into interactive 3D books, allowing readers to view and interact with content using AR and VR compatible devices.
Your published article is automatically converted into a realistic 3D book. Flip through pages and read research papers in a more engaging and interactive format.
Our website is actively being updated, and changes may occur frequently. Please clear your browser cache if needed. For feedback or error reporting, please email [email protected]
Thank you for connecting with us. We will respond to you shortly.
Lorem ipsum dolor sit amet, consectetur adipiscing elit. Ut elit tellus, luctus nec ullamcorper mattis, pulvinar dapibus leo.
New Representative of the Family Hydroscaphidae from Tunguska Basin, the Boundary of the Permian and Triassic (Coleoptera, Myxophaga) with Review on Myxophagan Fossil R ecords
Alexander G. Kirejtshuk<p>Russian Academy of Sciences</p>