Yam (Dioscorea spp.) is a vegetable of great economic and social importance, cultivated in tropical and subtropical regions. It is a staple food for millions of people, especially in West Africa, valued for its tuberous roots rich in carbohydrates, proteins, vitamins and minerals. In addition to its nutritional value, yam has cultural and medicinal significance, used in ceremonies and therapies. In Angola, climatic and geographic diversity favors the cultivation of different yam genotypes, resulting in a genetic heritage that is crucial for crop resilience and food security. The morphological characterization of genotypes allows the identification of varieties with desirable attributes, such as disease resistance, productivity and nutritional quality, facilitating genetic improvement programs. This study aims to morphologically characterize yam genotypes from different geographic origins in Angola, contributing to the knowledge and conservation of genetic diversity and the development of sustainable agricultural strategies.
## I. INTRODUCTION
Yam (Dioscoreaspp.) is a vegetable of great economic and social importance, widely cultivated in tropical and subtropical regions (Wang et al., 2023). It stands out as a staple food for millions of people, especially in West Africa, where species such as Dioscorea alata L. and Dioscorea rotundata (P.) JM are cultivated for their tuberous roots rich in carbohydrates, proteins, vitamins and minerals (Cazé Filho, 2002; Karkuteet al., 2017; In addition to its nutritional value, yam has cultural and medicinal significance, being used in various ceremonies and for therapeutic purposes (Padhan; Panda, 2020; Sousa; Raizada, 2020).
In Angola, with its varied climatic and geographical conditions, it offers a favorable environment for the cultivation of different yam genotypes (Kumar et al., 2014). The genetic diversity found in the country's yam plantations is the result of traditional agricultural practices, which involve the selection and propagation of local varieties adapted to the specific conditions of each region (Danquah et al., 2022). This genetic heritage is fundamental to the resilience of crops and the country's food security.
The morphological characterization of yam genotypes allows the identification of varieties with desirable attributes, such as disease resistance, productivity and nutritional quality, in addition to facilitating genetic improvement programs (Agre et al., 2019; Syombua et al., 2021; Norman et al. al., 2022).
To fully exploit the agricultural and economic potential of yam in Angola, it is crucial to obtain a detailed understanding of the morphological characteristics of the different genotypes present in the country. In this sense, this study aims to morphologically characterize yam genotypes from different geographic origins in Angola, aiming to contribute to the knowledge and conservation of the crop's genetic diversity, as well
as to the development of sustainable and efficient agricultural strategies.
## II. MATERIAL AND METHODS
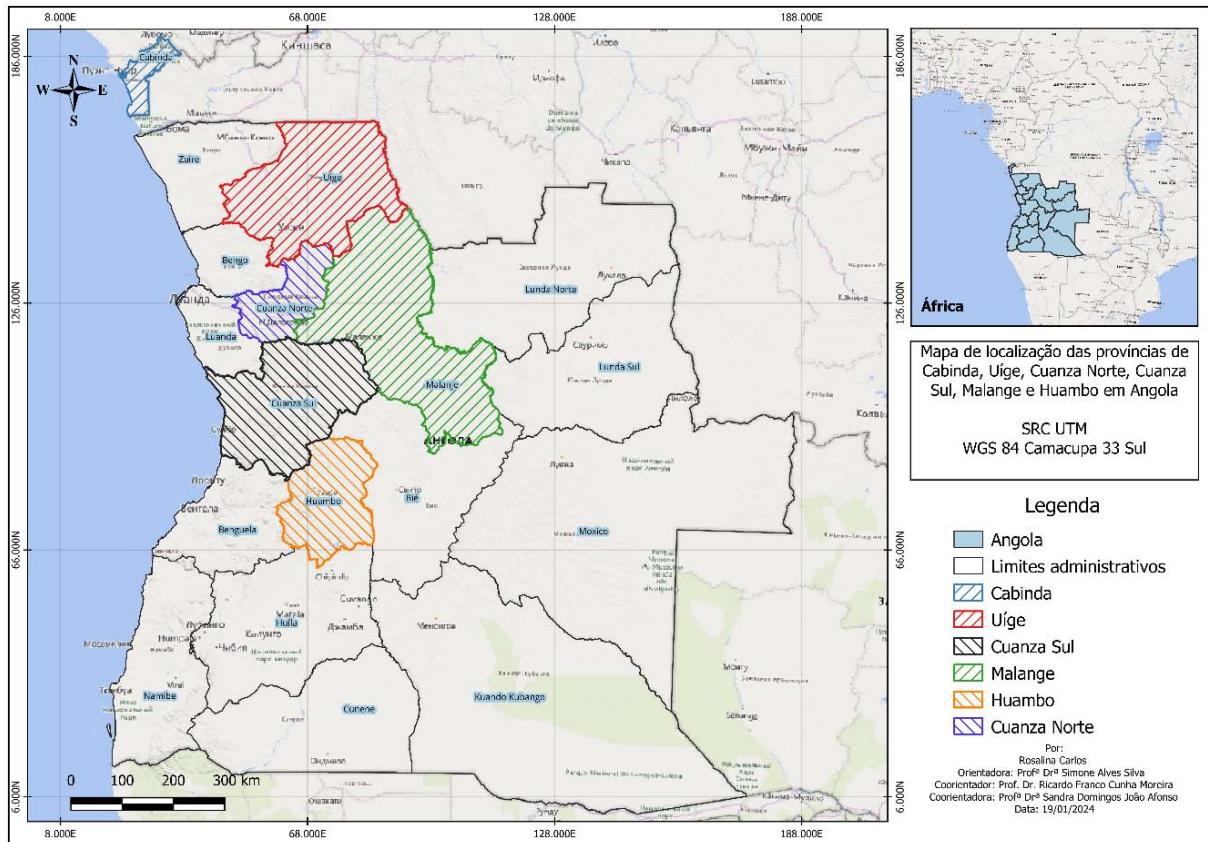
a) Obtaining and Processing Plant Material The accessions were collected from expeditions in the provinces of Cabinda, Cuanza Norte, Cuanza Sul,
Malange, Uige and Huambo, belonging to the country of Angola (Figure 1). Of these, we obtained two accessions Dioscorea esculenta (L.) B., five from Dioscorea alata L., six from Dioscorea Cayenesis L. and two from Dioscorea bulbifera L., as shown in Table 1.
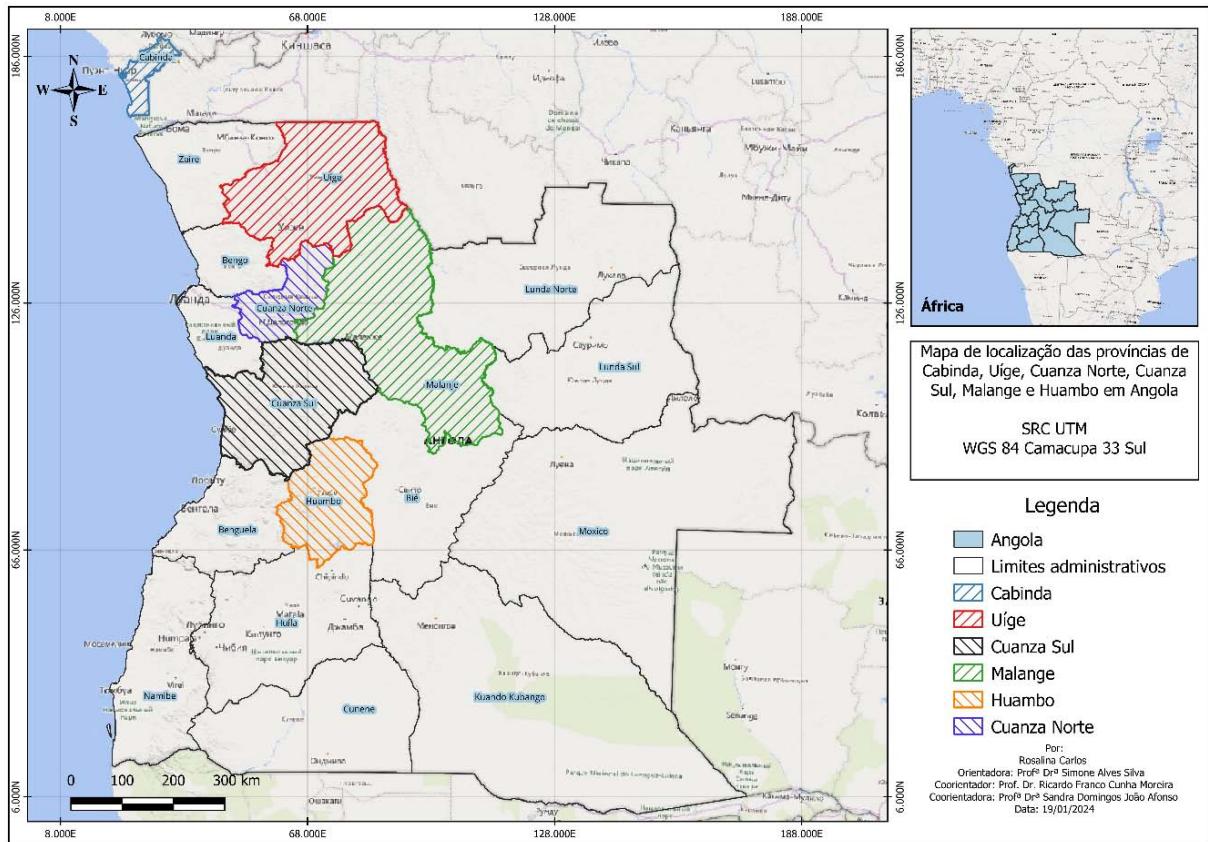
Figure 1: Location of yam (Dioscorea spp.) collection points in six provinces of Angola
Table 1: Geographic location of the yam accessions Dioscorea spp. in different provinces of Angola
<table><tr><td>Identification</td><td>Access</td><td>Common name</td><td>Geographic coordinates</td><td>Province</td></tr><tr><td>1</td><td>Dioscoreaesculenta</td><td>Assipi</td><td>12°46'48"S, 15°45'00"E and 1,700m</td><td>Huambo</td></tr><tr><td>two</td><td>Dioscoreaesculenta</td><td>Assipi</td><td>11°12'36"S, 13°50'24"E and 0 to 10m</td><td>Cuanza Sul</td></tr><tr><td>3</td><td>Dioscorea alata</td><td>Purple Mbanza</td><td>07°36'36"S, 15°03'00"E and 850m</td><td>Uíge</td></tr><tr><td>4</td><td>Dioscorea alata</td><td>N'guenge</td><td>09°18'00"S, 14°54'36"E and 1,180m</td><td>Cuanza Norte</td></tr><tr><td>5</td><td>Dioscorea alata</td><td>N'guenge</td><td>09°32'24"S, 16°20'24"E and 1,180m</td><td>Malange</td></tr><tr><td>6</td><td>Dioscorea alata</td><td>Mbambi</td><td>05°32'60"S, 12°11'60"E and 20m</td><td>Cabinda</td></tr><tr><td>7</td><td>Dioscorea alata</td><td>Yam</td><td>11°12'36"S, 13°50'24"E and 0 to 10m</td><td>Cuanza Sul</td></tr><tr><td>8</td><td>Dioscorea cayenesis</td><td>Yam</td><td>12°46'48"S, 15°45'00"E and 1,700m</td><td>Huambo</td></tr><tr><td>9</td><td>Dioscorea cayenesis</td><td>Yam potato</td><td>07°36'36"S, 15°03'00"E and 850m</td><td>Uíge</td></tr><tr><td>10</td><td>Dioscorea cayenesis</td><td>Mbambi</td><td>05°32'60"S, 12°11'60"E and 20m</td><td>Cabinda</td></tr><tr><td>11</td><td>Dioscorea cayenesis</td><td>Ngame</td><td>09°18'00"S, 14°54'36"E and 1,050m</td><td>North Cuanza</td></tr><tr><td>12</td><td>Dioscorea cayenesis</td><td>Yam</td><td>11°12'36"S, 13°50'24"E and 0 to 10m</td><td>Cuanza Sul</td></tr><tr><td>13</td><td>Dioscorea cayenesis</td><td>N'guenge</td><td>09°32'24"S, 16°20'24"E and 1,180m</td><td>Malange</td></tr><tr><td>14</td><td>Dioscorea bulbifera</td><td>Heart Potato</td><td>09°18'00"S, 14°54'36"E and 1,050m</td><td>Cuanza Norte</td></tr><tr><td>15</td><td>Dioscorea bulbifera</td><td>Heart Potato</td><td>07°36'36"S, 15°03'00"E and 850 m</td><td>Uíge</td></tr></table>
After collection, the materials were identified and documented with detailed information about each accession, which was obtained through interviews with
local producers, covering aspects such as local nomenclature, management methods and use of varieties.
### b) Area and Experimental Design
After obtaining access provided by rural producers, this material was taken to Fazenda Esperança, located in the city of Huíla (14°91'75"S, 13°49'25"E and 1,805m altitude), to be deployed in the field during the period from August 2022 to October 2023. The region is characterized by having a humid to dry subtropical climate, with an average annual temperature varying between $20^{\circ}\mathrm{C}$ and $22^{\circ}\mathrm{C}$. Precipitation is relatively uniform throughout the year, with a more intense rainy season from October to April, while the months of May to September tend to be drier.
The experimental design adopted was completely randomized, containing 15 replications of each species obtained, totaling a collection of 60 plants. Each genotype was arranged in plots consisting of three lines of 5 plants, totaling 15 plants per plot. Plants were spaced 1.0 to 1.2 meters between rows and 0.8 to 1.0
meters between plants within each row. During growth, management practices such as irrigation, staking and manual weeding were implemented. These practices ensured adequate plant development and minimized competition for resources.
### c) Morphological Characterization
After six months of planting, the characterization of the aerial part (leaves and stems) was carried out, using descriptors selected according to the key proposed by IPGRI/IITA (1997) for species of Dioscorea. 22 descriptors were evaluated, of which 15 were qualitative and quantitative (Table 2). The characterization of the aerial part was conducted with the aid of a ruler and analog universal caliper. The plants evaluated were those selected for tuber multiplication.
Table 2: Descriptors used for the morpho-agronomic characterization of yam accessions (Dioscoreaspp.) in Angola
<table><tr><td>Identification</td><td>Descriptor</td><td>Classes (codes)</td></tr><tr><td colspan="3">Stalk</td></tr><tr><td>1</td><td>Color</td><td>1. Green; 2. Green with purple bands; 3. Green with brown bands; 4. Purple</td></tr><tr><td rowspan="2">two</td><td>Wings</td><td>1. Gift; 2. Absent.</td></tr><tr><td>Wing color</td><td>1. green; 2. Purple</td></tr><tr><td>3</td><td>Thorns</td><td>1. Gift</td></tr><tr><td>4</td><td></td><td>2. Absent</td></tr><tr><td rowspan="2">5</td><td>Growth direction</td><td>1. Schedule; 2. Counterclockwise.</td></tr><tr><td>Presence of glasses</td><td>1. Gift; 2. Absent.</td></tr><tr><td>6</td><td>Diameter (15 cm from the base of the plant)</td><td>1. <0.4 cm; 2. 0.4 - 0.6 cm; 3. >0.6cm</td></tr><tr><td>7</td><td>Stem shape</td><td>1. Polygonal; 2. Round</td></tr><tr><td colspan="3">Sheets</td></tr><tr><td>8</td><td>Position</td><td>1. Alternate; 2. Opposite</td></tr><tr><td>9</td><td>Form</td><td>1. Cordata; 2. Sagitada</td></tr><tr><td>10</td><td>Petiole length</td><td>1. <5cm; 2. 5 - 10 cm; 3. >10 cm.</td></tr><tr><td rowspan="3">11</td><td>Petiole color</td><td>1. Green; 2. Green with brown; 3. Purple</td></tr><tr><td>Distance from the insertion of the petiole on the leaf to the upper end of the leaf (adult leaves)</td><td>1. <2cm; 2. 2 - 4 cm; 3. >4cm</td></tr><tr><td rowspan="2">Distance from the insertion of the petiole on the leaf to the lower end of the leaf (adult leaves)</td><td rowspan="2">1. <10cm; 2. 10 - 15 cm; 3. >15cm</td></tr><tr><td>13</td></tr><tr><td>14</td><td>Leaf width at largest portion (adult leaves)</td><td>1. <10 cm, 2. 10 - 15 cm; 3. >15cm</td></tr><tr><td colspan="3">Tubers</td></tr><tr><td>15</td><td>Underground</td><td>1. Gift; 2. Absent</td></tr><tr><td>16</td><td>Presence of aerial tubers</td><td>1. Gift; 2. Absent</td></tr><tr><td rowspan="2">17</td><td></td><td>1 one; 2. Some; 3. Many</td></tr><tr><td>Number of tubers</td><td>1. One; 2. Some; 3. Many</td></tr><tr><td>18</td><td>Form</td><td>1. Elongated; 2. Irregular; 3. Oval</td></tr><tr><td>19</td><td>Length</td><td>1. <20cm; 2. 20 - 40 cm; 3. >40cm</td></tr><tr><td>20</td><td>Width (major axis)</td><td>1. <7cm; 2. 7 - 12 cm; 3. >12cm</td></tr><tr><td>21</td><td>Shell color</td><td>1. Brown; 2. Yellow</td></tr><tr><td>22</td><td>Pulp color</td><td>1. White; 2. Yellow; 3. Purple; 4. Purple with white</td></tr></table>
At 11 months since implantation, the tubercles were characterized. The physical analyzes were carried out in the Laboratory of the Agricultural Research
Institute (IIA) of Huíla. The tubers were subjected to a visual inspection and evaluated for health, physical integrity, size and shape. They were then washed in
running water. Subsequently, they were weighed on a semi-analytical scale, peeled and sliced mechanically.
The measurements taken included the weight of the tuber with the skin (kg) and the weight of the tuber without the shell (g), which were obtained using a digital scale. In addition, the tuber length (cm), longitudinal and transverse diameter (cm), and longitudinal and transverse thickness (cm) were recorded. These measurements were carried out with the aid of a ruler and digital caliper. Subsequently, the tubers were cut in half and evaluated for longitudinal and transverse thickness. The weight of the tuber without the shell was measured again for recording.
### d) Statistical Analysis
For the set of data obtained through the descriptors, the percentage frequencies of each class and the entropy level of the descriptors were calculated using Renyi's entropy coefficient (Renyi, 1961). The physical characterization of the tubers was submitted to analysis of variance (ANOVA), using the F test, and the
means were compared using the Tukey test $(p < 0.05)$. Data analysis was performed using the R program (R Core Team, 2024).
## III. RESULTS AND DISCUSSION
Analysis of morphological data from yam accessions in several provinces of Angola revealed significant variability in the characteristics of the plants studied (Table 3). Among the 60 accessions analyzed, all had alternate leaves, suggesting a peculiar adaptation of these plants to the Angolan environment. The presence of alternate leaves in yam plants may be a reflection of their chromosomal evolution and genetic basis, as discussed by Bredeson et al. (2022). This characteristic may be related to agronomically important traits, as mentioned by Maroya et al. (2022), which address the transformation of yam seed systems in West Africa, highlighting the importance of producing high-quality seed tubers.
Table 3: Absolute, percentage and relative frequencies and entropy level for the evaluated descriptors of the Dioscoreaspp yam accessions. In different provinces of Angola
<table><tr><td>Descriptors</td><td>Classes</td><td>Frequency Absolute</td><td>Frequency Percentage (%)</td><td>Frequency Relative</td><td>Level of Entropy(H')</td></tr><tr><td rowspan="2">Sheet position</td><td>Alternate</td><td>60</td><td>100</td><td>1</td><td>0</td></tr><tr><td>Opposite</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="3">Leaf shapes</td><td>Cropped</td><td>15</td><td>25</td><td>0.25</td><td></td></tr><tr><td>Sagittadic</td><td>30</td><td>50</td><td>0.5</td><td>1.5</td></tr><tr><td>lobada</td><td>15</td><td>25</td><td>0.25</td><td></td></tr><tr><td rowspan="3">Number of leaf lobes</td><td>Absence</td><td>45</td><td>75</td><td>0.75</td><td></td></tr><tr><td>One</td><td>15</td><td>25</td><td>0.25</td><td>0.81</td></tr><tr><td>Three</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="3">Petiole length</td><td>< 5cm</td><td>13</td><td>21.67</td><td>0.22</td><td></td></tr><tr><td>5 - 10 cm</td><td>39</td><td>65</td><td>0.65</td><td>1.27</td></tr><tr><td>> 10 cm</td><td>8</td><td>13.33</td><td>0.13</td><td></td></tr><tr><td rowspan="3">Petiole color</td><td>Green</td><td>39</td><td>65</td><td>0.65</td><td></td></tr><tr><td>Green with brown</td><td>6</td><td>10</td><td>0.1</td><td>1.24</td></tr><tr><td>Purple</td><td>15</td><td>25</td><td>0.25</td><td></td></tr><tr><td rowspan="3">Distance from the insertion of the petiole on the leaf to the upper end</td><td>< 2cm</td><td>5</td><td>8.33</td><td>0.08</td><td></td></tr><tr><td>2 - 4 cm</td><td>18</td><td>30</td><td>0.3</td><td>1.25</td></tr><tr><td>> 4 cm</td><td>37</td><td>61.67</td><td>0.62</td><td></td></tr><tr><td rowspan="3">Distance from the insertion of the petiole on the leaf to the lower end</td><td>10 cm</td><td>26</td><td>43.33</td><td>0.43</td><td></td></tr><tr><td>10 - 15 cm</td><td>21</td><td>53</td><td>0.35</td><td>1.53</td></tr><tr><td>> 15 cm</td><td>13</td><td>21.67</td><td>0.22</td><td></td></tr><tr><td rowspan="3">Sheet width</td><td>< 10 cm</td><td>31</td><td>51.67</td><td>0.52</td><td></td></tr><tr><td>10 - 15 cm</td><td>29</td><td>48.33</td><td>0.48</td><td>1</td></tr><tr><td>> 15cm</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="3">Width between lobes</td><td>> 6 cm</td><td>45</td><td>75</td><td>0.75</td><td></td></tr><tr><td>6 - 10 cm</td><td>15</td><td>25</td><td>0.25</td><td>0.81</td></tr><tr><td>> 10 cm</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="4">Stem color</td><td>Green</td><td>45</td><td>75</td><td>0.75</td><td></td></tr><tr><td>Green with Purple strip</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td>Green with Brown band Purple</td><td>15</td><td>25</td><td>0.25</td><td>0.81</td></tr><tr><td></td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="2">Presence of thorn</td><td>Gift</td><td>15</td><td>25</td><td>0.25</td><td rowspan="2">0.81</td></tr><tr><td>Absence</td><td>45</td><td>75</td><td>0.75</td></tr><tr><td rowspan="2">Presence of wings</td><td>Gift</td><td>30</td><td>50</td><td>0.5</td><td rowspan="2">1</td></tr><tr><td>Absence</td><td>30</td><td>50</td><td>0.5</td></tr><tr><td rowspan="2">Wing color</td><td>Green</td><td>30</td><td>50</td><td>0.5</td><td rowspan="2">1</td></tr><tr><td>Purple</td><td>30</td><td>50</td><td>0.5</td></tr><tr><td rowspan="2">Presence of aculei</td><td>Gift</td><td>15</td><td>25</td><td>0.25</td><td rowspan="2">0.81</td></tr><tr><td>Absence</td><td>45</td><td>75</td><td>0.75</td></tr><tr><td rowspan="2">Growth direction</td><td>Time</td><td>15</td><td>25</td><td>0.25</td><td rowspan="2">0.81</td></tr><tr><td>Counter-clockwise</td><td>45</td><td>75</td><td>0.75</td></tr><tr><td rowspan="3">Stem diameter</td><td>< 0.4 cm</td><td>60</td><td>100</td><td>1</td><td></td></tr><tr><td>0.4 - 0.6 cm</td><td>-</td><td>-</td><td>-</td><td>0</td></tr><tr><td>> 0.6 cm</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="2">Stem shape</td><td>Polygonal</td><td>-</td><td>-</td><td>-</td><td rowspan="2">0</td></tr><tr><td>Round</td><td>60</td><td>100</td><td>1</td></tr><tr><td rowspan="2">Underground tubers</td><td>Gift</td><td>43</td><td>71.67</td><td>0.72</td><td rowspan="2">0.86</td></tr><tr><td>Absent</td><td>17</td><td>28.33</td><td>0.28</td></tr><tr><td rowspan="3">Number of tubers</td><td>One</td><td>6</td><td>10</td><td>0.1</td><td></td></tr><tr><td>Some</td><td>43</td><td>71.67</td><td>0.72</td><td>1.13</td></tr><tr><td>Many</td><td>11</td><td>18.33</td><td>0.18</td><td></td></tr><tr><td rowspan="3">Tuber shape</td><td>Elongated</td><td>19</td><td>31.67</td><td>0.32</td><td></td></tr><tr><td>Oval</td><td>26</td><td>43.33</td><td>0.43</td><td>1.55</td></tr><tr><td>Irregular</td><td>15</td><td>25</td><td>0.25</td><td></td></tr><tr><td rowspan="3">Tuber length</td><td>< 20 cm</td><td>46</td><td>76.67</td><td>0.77</td><td></td></tr><tr><td>20 - 40 cm</td><td>14</td><td>23.33</td><td>0.23</td><td>0.78</td></tr><tr><td>> 40 cm</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td rowspan="3">Tuber width</td><td>< 7 cm</td><td>28</td><td>46.67</td><td>0.47</td><td></td></tr><tr><td>7 - 12 cm</td><td>24</td><td>40</td><td>0.4</td><td>1.43</td></tr><tr><td>> 40cm</td><td>8</td><td>13.33</td><td>0.13</td><td></td></tr><tr><td rowspan="3">Shell color</td><td>Brown</td><td>45</td><td>75</td><td>0.75</td><td></td></tr><tr><td>Yellow</td><td>-</td><td>-</td><td>-</td><td>0.81</td></tr><tr><td>Purple</td><td>15</td><td>25</td><td>0.25</td><td></td></tr><tr><td rowspan="5">Tuber color</td><td>White</td><td>46</td><td>76.67</td><td>0.77</td><td></td></tr><tr><td>Yellow</td><td>14</td><td>23.33</td><td>0.23</td><td></td></tr><tr><td>Purple</td><td>-</td><td>-</td><td>-</td><td>0.78</td></tr><tr><td>Purple with white</td><td>-</td><td>-</td><td>-</td><td></td></tr><tr><td>White with purple</td><td>-</td><td>-</td><td>-</td><td></td></tr></table>
Regarding the shapes of the leaves, $50\%$ of the accessions had sagitate leaves, $25\%$ had lobed leaves, and another $25\%$ had cut leaves. The sagittal shape of the leaves plays a crucial role in the efficiency of light capture and water drainage. This format is advantageous in environments with high humidity, such as some areas of Angola, where better drainage can minimize the proliferation of pathogens (Norman; Tongoona; Shanahan, 2011). According to Ferriol et al. (2023), the presence of sagitate leaves can be seen as an adaptive strategy to maximize sunlight capture, reflecting an evolutionary response to local environmental conditions.
In addition to the shape of the leaves, it was observed that the majority of accessions (75%) did not have lobes, while 25% exhibited one lobe. The absence of lobes may be related to greater efficiency in regulating water loss through transpiration and optimization of light capture, as indicated by Brodribb et al. (2007). Unlobed leaves can facilitate a smaller
exposed surface area, reducing evapotranspiration and potentially improving water retention in environments where water availability is a critical constraint.
In terms of petiole length, $65\%$ of accessions had petioles between 5 and $10~\mathrm{cm}$ in size, suggesting a possible morphological adaptation to optimize the capture of environmental resources. The predominant color of the petioles was green $(65\%)$, which suggests an efficient photosynthetic capacity. The green color of the petioles indicates a high density of chlorophyll, which contributes to the efficiency of photosynthesis, especially under variable light conditions (Riekotte et al., 2022). This adaptation is essential to ensure that the plant can maximize light absorption and, consequently, energy production in different lighting scenarios.
Analysis of the distance between the insertion of the petiole on the leaf and the upper end revealed that $61.67\%$ of the accessions had a distance greater than 4 cm. This characteristic may indicate a strategy to maximize exposure to sunlight, allowing the leaves to be
arranged to avoid mutual shading and ensuring better light capture (Onwueme; Johnston, 2000). This arrangement can also influence photosynthesis efficiency and biomass production, reflecting an adaptive response to different light conditions.
Regarding the width of the leaves, $51.67\%$ of the accessions had leaves with a width of less than 10 cm, while $75\%$ showed a width between lobes greater than 6 cm. Variation in leaf width and distance between lobes can directly affect hydraulic conductivity and maximum leaf photosynthesis rate, as discussed by Tsukaya (2006). These morphological differences can influence the adaptation of plants to different environments and their ability to compete for resources.
For stem color, $75\%$ of accessions had green stems, while $25\%$ displayed stems with a brown band. The green color of the stem is associated with the presence of chlorophyll, which may contribute to supplementary photosynthesis, while stems with brown bands may indicate greater lignification or the presence of defense compounds (Aremuet al., 2019). These characteristics may reflect adaptations to environmental pressures, such as the need for defense against herbivores or the optimization of photosynthetic efficiency.
The morphological diversity observed in yam plants, particularly in leaf shapes, petioles and stem colors, indicates the plants' varied strategies for optimizing light capture and resource utilization, highlighting the importance of genetic and morphological diversity in increasing adaptability of plants to specific environments. Studies in Tanzania and Uganda have characterized yam genotypes based on leaf characteristics, flowering characteristics, and tuber shapes, revealing substantial phenotypic variability within the species (Massawe; Temu, 2022; Ferriol, 2023).
The presence of thorns was observed in $25\%$ of the accessions, possibly as an adaptation for defense against herbivores. Spines are defense mechanisms that make it difficult for herbivores to access tubers, reducing predation and helping the plant survive in environments where herbivore pressure is high (Nilsen; Orcutt, 1996). Such adaptation may be essential for the preservation of energy reserves in tubers, ensuring the availability of resources for growth and reproduction in adverse conditions.
The growth direction was predominantly counterclockwise (75%), and all stems had a round shape. The counterclockwise growth direction observed in 75% of accessions may have implications for the architecture of the root and tuber system. This orientation may be related to internal growth mechanisms that favor a more efficient distribution of tubers in the soil, optimizing the absorption of nutrients and water (Khan; Gement; Villordon, 2016). Such a growth pattern can also minimize competition between
roots and tubers, allowing for better development and more efficient use of available resources.
In relation to underground tubers, $71.67\%$ of the accessions had tubers, with the majority having one or a few tubers, and only $18.33\%$ having many tubers. Tubercle shapes varied, with $31.67\%$ elongated, $43.33\%$ oval and $25\%$ irregular. Variability in tuber shapes has a significant impact on storage capacity and resource use efficiency. Elongated tubers, which correspond to $31.67\%$ of accessions, offer a greater contact surface with the soil, which can facilitate nutrient absorption. Oval and irregular tubers can adapt better to different types of soil and environmental conditions, promoting a more uniform distribution of nutrients and better adaptation to variations in the environment (Epping; Laibach, 2020). This morphological diversity may reflect adaptive strategies to optimize resource use efficiency and maximize survival in different ecological conditions.
The majority of tubers were less than 20 cm long (76.67%) and less than 7 cm wide (46.67%), which may indicate an adaptive strategy to maximize biomass production and agricultural yield. Smaller tubers tend to be more efficient in terms of growth, allowing for greater planting density and better use of available space and resources (Squire, 2004). They can also facilitate harvesting and reduce processing costs, making production more efficient and economically viable.
As for the color of the shell, $75\%$ were brown and $25\%$ purple. These characteristics may reflect adaptations to different environmental conditions and abiotic stresses. Tubers with brown skin, observed in $75\%$ of accessions, are generally more resistant to physical damage and pathogen attacks due to the presence of phenolic compounds that act as natural barriers (Lattanzio et al., 2006). Tubers with purple skin, which represent $25\%$ of accessions, contain anthocyanins, which protect against oxidative stress and UV radiation, contributing to the durability and resistance of plants in adverse environments.
The color of the tuber was predominantly white (76.67%), followed by yellow (23.33%). The color of the tuber pulp has direct implications for the nutritional value and acceptability of the final product. White pulps, predominant in 76.67% of accessions, are typically rich in starch and provide an effective source of energy (Ukwuru; Egbonu, 2013). The yellow pulp, present in 23.33% of accessions, may indicate the presence of carotenoids, which are important for human health as precursors of vitamin A. The color of the pulp also influences consumer preference, affecting demand and value market for tubers.
Entropy was used to measure the variability of each descriptor, where higher values indicate greater diversity and lower values indicate homogeneity (Jost, 2006). All accessions had alternate leaves, stems with a diameter of less than $0.4\mathrm{cm}$, and round stems. This
homogeneity may be the result of natural or artificial selection that favored specific characteristics, adapted to the local environment or prevalent agricultural practices (Muraleedharan; Rajan, 2024). The complete absence of variability may, in some cases, indicate successful adaptation to stable environmental conditions, but it may also limit the ability of plants to respond to environmental changes or new biotic challenges (Vishnu; Rajan; Jaishanker, 2023).
In contrast, some descriptors exhibit high entropy, such as leaf shapes ( $H' = 1.5$ ) and growth direction ( $H' = 1.55$ ). The shapes of the leaves show an almost uniform distribution between cut (25%), sagitate (50%) and lobed (25%), while the direction of growth presents a balanced distribution between clockwise and counterclockwise growth. High entropy for these descriptors indicates high genetic variability, suggesting the presence of multiple growth forms and strategies within the population. This may be beneficial for adaptation to different environmental conditions and selection pressures. It would be useful to review articles that discuss the importance of genetic variability in adapting plants to variable environments and how this can influence survival and productivity.
Descriptors with moderate entropy, such as the number of leaf lobes $(\mathsf{H}^{\prime} = 0.81)$ and the presence of spines $(\mathsf{H}^{\prime} = 0.81)$, indicate that, although there is a predominant characteristic, there is a notable presence of variability. Most accessions do not have lobes (75%), but 25% have a lobe. Similarly, the predominance of accessions without thorns (75%) is accompanied by 25% of accessions with thorns. The moderate entropy suggests that these characteristics are important for adaptation and may vary according to different selective pressures. Furthermore, studies on yam genotypes in Tanzania revealed high morphological variabilities among 74 genotypes, with characteristics such as thorns at the base of the stem contributing to the observed variabilities (Massawe; Temur, 2022). This variability in morphological characteristics is crucial for understanding the genetic diversity and possible
adaptations of yam species, highlighting the importance of these characteristics in breeding programs and yam selection processes (IAdejumobi et al., 2023).
It is relevant to investigate how the presence or absence of certain morphological traits, such as lobes and spines, can affect the plant's interaction with its environment and its pathogens (Pérez-Harguindeguy et al., 2016).
Descriptors such as petiole length $(\mathsf{H}^{\prime} = 1.27)$, petiole color $(\mathsf{H}^{\prime} = 1.24)$ and wing color $(\mathsf{H}^{\prime} = 1.24)$ show a relatively balanced distribution between classes, indicating a significant phenotypic diversity that can be associated with genetic variations or environmental influences. Most petioles measure between $5 - 10~\mathrm{cm}$, with lower frequencies for other classes. The color of the petiole is predominantly green, but there are significant presences of purple and green petioles with brown. The color of the wings has a balanced distribution between green and purple. The balanced distribution of these descriptors suggests that multiple variables can influence the phenotypic expression of these traits, and it is important to understand how these variations can impact plant functionality and efficiency, allowing populations to persist and adapt to changing conditions (Lasky et al., 2023).
Finally, descriptors with low entropy, such as stem color $(H^{\prime} = 0.81)$ and the presence of spikes $(H^{\prime} = 0.81)$, indicate a clear predominance of certain characteristics, but with a slight presence of variability. Most stems are green, with a smaller presence of green stems with a brown band, and most accessions do not have needles. Low entropy suggests a clear predominance of certain characteristics, possibly indicating selection for more favorable characteristics (Vishnu; Rajan; Jaishanker, 2023).
analysis of the data in Table 4 reveals notable differences between yam species in relation to all morphometric characteristics evaluated. The extremely low p-values indicate a high statistical significance in these differences, highlighting the importance of these variables in differentiating between species.
Table 4: $p$ value, coefficient of variation (CV) and means for tuber and pulp weights (g), tuber length (cm), longitudinal and transverse diameters (mm) and transverse and longitudinal thicknesses (cm) of the accesses from yam Dioscoreaspp. In different provinces of Angola
<table><tr><td>Variables</td><td>p-value</td><td>CV (%)</td><td>Average General</td><td>Dioscorea the can</td><td>Dioscorea Cayenesis</td><td>Dioscorea bulbifera</td><td>Dioscorea esculent</td></tr><tr><td>Tuber weight</td><td>2.00 × 10-16</td><td>39.47</td><td>337.43</td><td>413.13b</td><td>143.87c</td><td>705.73a</td><td>87.00c</td></tr><tr><td>Pulp weight</td><td>1.84 × 10-14</td><td>54.3</td><td>259.17</td><td>269.53b</td><td>122.20c</td><td>588.53a</td><td>56.40c</td></tr><tr><td>Length</td><td>2.00 × 10-16</td><td>23.76</td><td>15.44</td><td>19.63b</td><td>8.66c</td><td>27.87a</td><td>5.59c</td></tr><tr><td>Longitudinal diameter</td><td>2.00 × 10-16</td><td>18.89</td><td>51.05</td><td>48.92b</td><td>33.38c</td><td>78.47a</td><td>43.43b</td></tr><tr><td>Transverse diameter</td><td>6.75 × 10-6</td><td>17.65</td><td>60.89</td><td>62.95ab</td><td>60.40b</td><td>71.40a</td><td>48.81c</td></tr><tr><td>Transverse thickness</td><td>2.00 × 10-16</td><td>14.98</td><td>5.13</td><td>5.41b</td><td>4.10c</td><td>7.11a</td><td>3.90c</td></tr><tr><td>Longitudinal thickness</td><td>2.00 × 10-16</td><td>12.77</td><td>5.52</td><td>4.95b</td><td>5.10b</td><td>7.89a</td><td>4.14c</td></tr></table>
The weight of the tuber (PT) and pulp (PP) showed considerable variation between species (Figure 2A and 2B respectively). Dioscorea bulbifera (DB) stood out significantly with substantially higher averages for both characteristics: $337.43\mathrm{g}$ for tuber weight and $259.17\mathrm{g}$ for pulp weight, compared to the other species. These high values suggest a superior energy storage capacity, which can be attributed to an
efficiency in the accumulation of reserves. The coefficients of variation (CV) indicate significant morphological diversity for these characteristics. In contrast, Dioscorea esculenta (DE) showed the lowest mean values for tuber and pulp weight, reflecting a different adaptive strategy, possibly favoring smaller and lighter tubers, as evidenced in the lower mean tuber weight values.
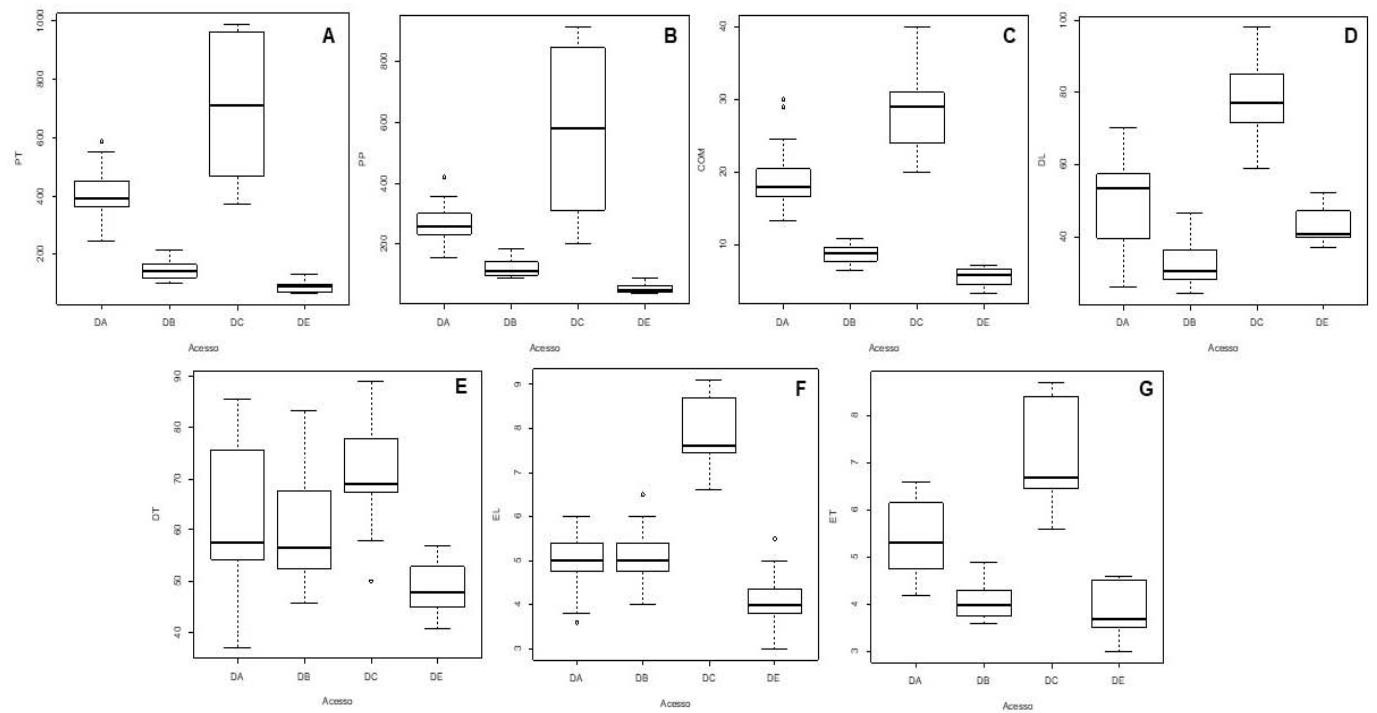
Figure 2: Averages for tuber (PT) and pulp (PP) weights, tuber length (COM), longitudinal (DL) and transverse (DT) diameters and transverse (ET) and longitudinal (EL) thicknesses for Dioscoreaaccessions alata (DA), Dioscoreacayenesis (DC), Dioscoreabulbifera (DB) and Dioscoreaesculenta (DE) in different provinces of Angola
Some Dioscorea bulbifera genotypes exhibited notably larger aerial tuber weights and diameters, which contributed to an increase in overall tuber production per plant. Genetic variability and heritability analyzes revealed significant differences between plant accessions in terms of yield and yield-related traits, with the potential for larger and heavier tubers in certain accessions. These findings suggest that the species has significantly larger and heavier tubers, highlighting the variability in tuber characteristics (Lu et al., 2024).
Furthermore, it was observed that Dioscorea bulbifera had significantly larger and heavier tubers, results similar to those found in Angola. The efficiency in accumulating energy reserves in this species may be an advantageous adaptive characteristic for biomass production in agricultural contexts (Rayamajhi et al., 2020).
For tuber length (COM) and longitudinal diameter (DL), Dioscorea bulbifera (DB) also presented the highest average values: 27.87 cm for length and 78.47 mm for longitudinal diameter. Dioscorea alata (DA) and Dioscorea cayenensis (DC) showed
intermediate values, while Dioscorea esculenta (DE) showed the lowest values, with an average of 5.59 cm for tuber length and 43.43 mm for longitudinal diameter (Figure 2C and 2D). The transverse diameter (DT) follows a similar pattern, with Dioscorea bulbifera (DB) again exhibiting the highest average values (71.40 mm), standing out for its ability to grow in volume (Figure 2E).
Regarding longitudinal (EL) and transverse (ET) thicknesses, Dioscorea Cayenensis (DC) and Dioscorea alata (DA) present higher values for transverse thickness, while Dioscorea bulbifera (DB) has the highest longitudinal thickness. Dioscorea esculenta (DE) exhibited the lowest thicknesses for both parameters, reinforcing the trend towards smaller and more compact tubercles (Figure 2F and 2G).
The work by Silva, Araújo Pires (2020) also reinforces these conclusions, highlighting that Dioscorea bulbifera showed high efficiency in storing carbohydrates, which is beneficial in agricultural systems that prioritize biomass production. The study suggests that the ecological adaptability of this species may explain its ability to produce larger and heavier
tubers. In contrast, Dioscorea esculenta tends to produce smaller tubercles, possibly due to an ecological adaptation to less fertile soils and more challenging environmental conditions.
The differences observed in morphometric and morphological characteristics between yam species indicate significant genetic diversity within the genus Dioscorea in Angola. Dioscorea bulbifera (DB) stands out for its larger and heavier tubercles, which suggests an efficient adaptation to store large amounts of energy reserves. This characteristic is especially valuable in agricultural contexts where biomass production is a priority. This capability can be beneficial for uses where tuber volume and weight are critical to yield.
On the other hand, species such as Dioscorea esculenta (DE), with smaller and lighter tubercles, may be adapted to specific environmental conditions or agricultural systems that favor smaller tubercles. These characteristics may reflect adaptations to different ecological niches or cultivation strategies, where smaller tubers may be advantageous.
Furthermore, Chen et al. (2022) discuss how genetic diversity among Dioscorea species can influence these morphological and adaptive variations. The study revealed that variations in longitudinal and transverse diameters, as well as transverse and longitudinal thicknesses, can influence not only crop yield, but also resistance to pests and diseases, storage capacity and nutritional quality. Therefore, when selecting yam varieties for cultivation, it is crucial to consider both morphometric characteristics and environmental requirements and cultivation preferences, to maximize the yield and efficiency of agricultural production.
## IV. CONCLUSION
There is significant morphological and morphometric variability among yam accessions in the provinces of Angola. Dioscorea bulbifera stood out for the greater weight and dimensions of its tubes, indicating a greater nutrient storage capacity and structural robustness, in contrast to Dioscorea aesculenta and Dioscorea cayenesis, which presented smaller measurements.
Generating HTML Viewer...
Funding
No external funding was declared for this work.
Conflict of Interest
The authors declare no conflict of interest.
Ethical Approval
No ethics committee approval was required for this article type.
Data Availability
Not applicable for this article.
How to Cite This Article
Rosalina Esperança da Silva Carlos. 2026. \u201cMorphological Characterization of Yam Genotypes from Different Geographic Origins in Angola\u201d. Global Journal of Science Frontier Research - D: Agriculture & Veterinary GJSFR-D Volume 24 (GJSFR Volume 24 Issue D1).
Explore published articles in an immersive Augmented Reality environment. Our platform converts research papers into interactive 3D books, allowing readers to view and interact with content using AR and VR compatible devices.
Your published article is automatically converted into a realistic 3D book. Flip through pages and read research papers in a more engaging and interactive format.
Our website is actively being updated, and changes may occur frequently. Please clear your browser cache if needed. For feedback or error reporting, please email [email protected]
Thank you for connecting with us. We will respond to you shortly.