Otolith morphology analysis is one of the main tools used for fish or stock identification. Moreover, otolith shape can also be used in the fish feeding studies (stomach content) for the identification of prey fish and their size according to the relationship between fish and otolith sizes. In the present study, the relationship between fish length and otolith morphological dimensions was investigated for Sargocentron spiniferum (Forsskål, 1775) (family: Holocentridae). The samples (185 fish and 370 sagittal otoliths) were collected from the coast of the Red Sea, Egypt. The statistical analysis was undertaken in two steps using generalized linear models for the relationship between body length and weight and otolith morphology descriptors (length, width, area, and perimeter) and shape indices (aspect ratio, compactness, form factor, rectangularity, roundness, ellipticity, squareness, sulcus, and ostium). From the relationships between total length (TL) of fish and fourteen morphology descriptors and shape indices, three are significantly correlated with TL (otolith length, cauda, and squareness) where the side effect were p
## I. INTRODUCTION
The sabre squirrelfish Sargocentron spiniferum (Forsskal, 1775) is a member of the family Holocentridae, which is mainly distributed in the Indo-Pacific from the Red Sea and East Africa to the Hawaiian Islands and Ducie Islands extending south to Australia. It is also distributed in New Caledonia, north to southern Japan and the Ogasawara Islands (Randall, 1998; Randall and Greenfield, 1999). This species inhabits different reef areas between reef flats in lagoons and seaward reefs at a depth of $122\mathrm{m}$. This fish occurs under ledges during the day (Lieske and Myers, 1994), and when smaller in size, this fish inhabits shallow protected areas. It is a nocturnal fish that feeds on crabs, shrimp, and small fishes (Kuiter and Tonozuka, 2001).
Many researchers have been focusing on the study age and growth of fishes due to the clear and distinct growth rings of sagittal otoliths. Otolith has calcified structures and located at the right and left inner ears of fishes and it is useful in the study of fish biology, ecology, and fisheries science (Lecomte-Finiger, 1999; Tuset et al., 2003; Jawad et al., 2017; Mehanna et al., 2019; Serdar and Derya, 2020). Also, the otoliths are used to estimate movement, varied habitat, population dynamics, and trophic ecological level of ecosystem for fish species (Campana and Casselman, 1993; Tuset et al., 2003a; Cardinale et al., 2004; Rooker et al., 2008; Zorica et al., 2010; Morat et al., 2012; Yilmaz et al., 2014; Osman et al., 2020).
Otolith shape and dimension are used to identify fish species or stocks. However, a recent study showed that the directional asymmetry between right and left otoliths within individuals could affected the results from the otolith shape analysis as tool to identify the stocks (Mahé et al., 2018b). So, the aim of the present work was to estimate the relationship between fish size and otolith size (length, width, weight, form factor, and aspect ratio) to identify this species from the Egyptian water of the Red Sea.
## II. MATERIALS AND METHODS
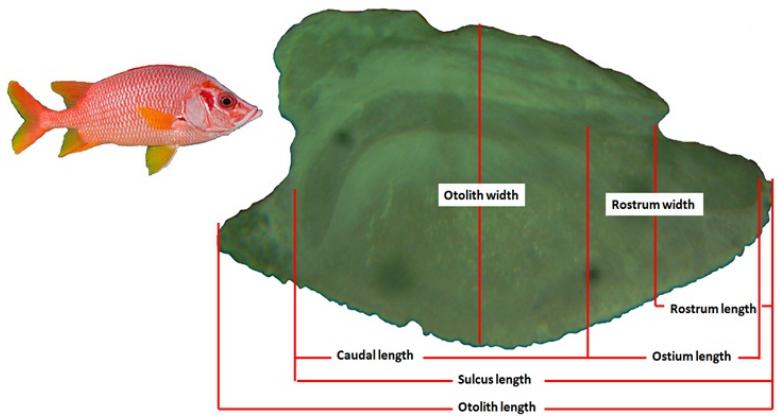
Fish species were randomly collected from the southern Red Sea at the Shalateen fishing port, which is located 520 km south of Hurghada (Fig. 1), Egypt, during March 2018 to February 2019. The fish were obtained from the commercial catch of the hook and line fishery at Shalateen fishing ground. In the laboratory, total fish length (TL) was measured to the nearest 0.1 mm, and fish weight (W) was recorded to the nearest 0.01 g. Then, the sex was determined. The total length and body weight of the species ranged between 17.7 and 45.8 cm and 101.5 and 1632 g respectively. Sagittal otoliths (370 left and right otoliths) were extracted from the inner ear of 185 S. spiniferum, cleaned and dried. Otolith weight (OW) for each head side was measured using a digital balance AS220 k/1 to the nearest 0.0001 g. Otolith outlines were realised using a Euromex-CMEX- 10 PRO camera with a stereomicroscopic. Otolith length (OL, mm), otolith area (OA, mm2), otolith perimeter (OP, mm), sulcus (SU), ostuim (OS), Form factor (FF), Aspect ratio (AR), circularity (CI) rectangularity (RE), Round (RO), Ellipticity (EL), Compactness (C), and Squaresness (SQ) were extracted using Image J analysis software (Rohlf, 2006) (detailed descriptions is in table 1 and Figure 2). The shape index factors (FF, C, AR, CI, RO, RE, EL, and SQ) were calculated according to different formulae (Table 1) (Russ, 1990; Tuset et al., 2003b; Pavlov et al., 2015; Mahe et al., 2016; Zischke et al., 2016; Jawad et al.,
2017; Kabakli and Ergüden, 2018; Mahé et al., 2018a; Osman et al., 2020). The main step was modelled the relationship of body length with the otolith outline indices according to the side (S):
$$
\log LT \sim OL + OW + OH + OA + OP + SU + OS + FF + AR + CI + RE + RO + EL + C + SQ + OL:S + OH:S + OW:S + OA:S + OP:S + SU:S + OS:S + FF:S + AR:S + C:S + SQ
$$
Statistical analyses were performed in the statistical environment R (Fox and Weisberg, 2011) stats package (R Development Core Team 2016).
The otoliths of S. spiniferum, shape of the sulcus acusticus, sulcus type, ostium, the caudal, dorsal and ventral marginal have similar descriptions, and the characteristics of an otolith of S. spiniferum were detailed descriptions are in Table 2). A general pattern of S. spiniferum sagittae can be recognized in adult individuals.
Otolith shape of S. spiniferum is ovate with sinuate margins and the otolith is very thick. The sulcus acousticus is ostial with the Heterosulcus and ostium formed by a short funnel-like ostium that opens to the anterior margin and closed tubular cauda at least two times larger than the ostium (Figure 2 and Table 2). The statistical description and paired t-test results for left and right otoliths of S. spiniferum are given inTable 3.
The analysis of the relationship between fish length and fourteen otolith shape descriptors using a generalized linear model showed that there is a significant relationship between eight otolith parameters (ostium, cauda, otolith area, otolith perimeter, compactness, form factor, circularity, and squareeness), with the total length of fish. Only the relationship of body length with the otolith length, cauda and squareness $(P < 0.05)$ was significant for right and left otolith (Table 4).
The correlation between fish total length and otolith morphology showed that with the increase of the total length, the otolith morphology such as otolith length, width, sulcus, ostum cauda, area, and perimeter also increase (Figure 3). These relationships between body length and otolith measurements (left and right) were best fitted as linear regression. On the other hand, the relationship between body length and the aspect ratio was close to 1; confirming that the otolith of S. spiniferum was oval. There is a positive relationship between total length and the aspect ratio. The same was noticed for the compactness and circularity values where they increased as the TL increased. The form factor, rectangularity, roundness, ellipticity, and squatness values decreased as the TL decreased.
## III. DISCUSSION
The sabre squirrelfish Sargocentron spiniferum (Forsskål, 1775) contribute to important fisheries, specially the small artisanal fisheries at Shalatin fishing ground, Red Sea, Egypt. Form this point, basic data on the biology and dynamics of the species are essential for successful stock assessment and consequently in fisheries management.
The observed fish length and shape of the otolith in this study should encourage more research to verify the essential role of otolith morphometric measurements in fish stock identification. The strong correlation between the somatic length and otolith size suggests that somatic growth has a significant influence on the otolith growth (Jockusch, 1997; Cardinale et al., 2004). The results of this study were not similar to those previously obtained for the redcoat S. rubrum (Kabakli and Ergüden, 2018), where the present study have max otolith length, weight and hight than S. rubrum. Previous studies have focused mainly on the relationship between otolith measurements and fish length (Harvey et al., 2000; Fossen et al., 2003; Lychakov et al., 2006; Morat et al., 2008; Pavlov, 2016; Mehanna et al., 2016; Osman et al., 2020). In our study, the results of generalized linear model showed the relationship among these parameters as the otolith length could be affected by the choice of the otolith (significant asymmetry between right and left otoliths). Based on the present data, the relationship between TL and AR, CO, and CI was determined as a linear relationship, despite the relationship among TL and FF, RE, RO, EL, and SQ being determined as a nonlinear relationship.
On the other hand, the shape of otolith from different geographical areas is influenced by abiotic ecological parameters (e.g. salinity, temperature) and biotic parameters for examples prey availability, and depends on individual genotype (Cardinale et al. 2004; Gagliano and McCormick, 2004; Swan and Palmer, 2006; Vignon and Morat, 2010). So, an interaction of environmental and genetic fluctuation generates the morphological variance in shape of otolith that may allow the differentiation of stock units. However, the factors that affects the shapes are not fully understood and have not been investigated deeply yet (Burke et al., 2008). An on-going recent work displayed that the ontogenetic trajectory of otolith shape could be impacted by the environmental disturbance during early life stage (Vignon, 2018).
The relationships between fish size and otolith shape indices demonstrate the high variability in fish length and morphometric parameters, indicating that the otolith of S. spiniferum is rectangular to oval. As results in the current study, the fish size and otolith morphometric parameters are useful for further research on verifying the role of otoliths identification, discrimination and taxonomic classification of fish. Also, the results showed that the otolith shape indices significantly differed from species to species, although the indices indicate a similar pattern for otoliths. The results are reliable with that illustrated that otoliths are widely used for the discrimination and variation of fish species because of their form, diet, weight, and growth (Tuset et al., 2008; Bacha et al., 2010).
Finally, the estimation of the generalized linear model supposed in the present work may be good tool to study the relationship between fish and otolith morphometric features used to fish population dynamics, stomach contents of piscivorous predators, paleontological composition, and yield estimates.
## IV. CONCLUSION
Finally, the estimation of the generalized linear model supposed in the present work may be good tool to study the relationship between fish and otolith morphometric features used to fish population dynamics, stomach contents of piscivorous predators, paleontological composition, and yield estimates.
Generating HTML Viewer...
References
44 Cites in Article
M Bacha,R Amara (2010). Spatial, temporal and ontogenetic variation in diet of anchovy (Engraulis encrasicolus) on the Algerian coast (SW Mediterranean).
N Burke,D Broph,P King (2008). Otolith shape analysis: its application for discriminating between stocks of Irish Sea and Celtic Sea herring (Clupea harengus) in the Irish Sea.
Steven Campana,John Casselman (1993). Stock Discrimination Using Otolith Shape Analysis.
Steven Campana,John Neilson (1985). Microstructure of Fish Otoliths.
M Cardinale,P Doering-Arjes,M Kastowsky,H Mosegaard (2004). Effects of sex, stock, and environment on the shape of known-age Atlantic cod (<i>Gadus morhua</i>) otoliths.
I Fossen,O Albert,E Nilssen (2003). Improving the precision of ageing assessments for long rough dab by using digitised pictures and otolith measurements.
J Fox,S Weisberg (2011). An {R} Companion to Applied Regression.
R Froese (2006). Cube law, condition factor and weight-length relationships: history, meta-analysis and recommendations.
M Gagliano,M Mccormick (2004). Feeding history influences otolith shape in tropical fish.
J Harvey,T Loughlin,M Perez,D Oxman (2000). Relationship between fish size and otolith length for 63 species of fishes from the eastern North Pacific Ocean NOAA Tech Rep.
K Hüssy (2008). Otolith shape in juvenile cod (Gadus morhua): Ontogenetic and environmental effects.
Laith Jawad,Kristiaan Hoedemakers,Ana Ibáñez,Yassin Ahmed,Mohamed Abu El-Regal,Sahar Mehanna (2017). Morphology study of the otoliths of the parrotfish,<i>Chlorurus sordidus</i>(Forsskål, 1775) and<i>Hipposcarus harid</i>(Forsskål, 1775) from the Red Sea coast of Egypt (Family: Scaridae).
Elizabeth Jockusch (1997). Geographic Variation and Phenotypic Plasticity of Number of Trunk Vertebrae in Slender Salamanders, Batrachoseps (Caudata: Plethodontidae).
Ferhat Kabakli,Deniz Erguden (2018). Age, Growth, and Mortality of the Redcoat Sargocentron rubrum (Forsskal, 1775), in Iskenderun Bay, Northeastern Mediterranean.
R Kuiter,T Tonozuka (2001). Pictorial guide to Indonesian reef fishes.
Raymonde Lecomte-Finiger (1999). L'otolithe: la ≪ boîte noire ≫ des Téléostéens.
E Lieske,R Myers (1994). Coral reef fishes Caribbean, Indian Ocean and Pacific Ocean including the red sea.
D Lychakov,Y Rebane,A Lombarte,L Fuiman,A Takabayashi (2006). Fish otolith asymmetry: Morphometry and modeling.
K Mahé,Y Aumond,K Rabhi,R Elleboode,E Bellamy,J Huet,M Gault,D Roos (2017). Relationship between somatic growth and otolith growth: a case study of the ornate jobfish<i>Pristipomoides argyrogrammicus</i>from the coast of Réunion (SW Indian Ocean).
K Mahé,E Bellamy,J Delpech,C Lazard,M Salaun,Y Vérin,F Coppin,M Travers-Trolet (2018). Evidence of a relationship between weight and total length of marine fish in the Northeastern Atlantic Ocean: physiological, spatial and temporal variations.
K Mahe,H Evano,T Mille,D Muths,J Bourjea (2016). Otolith shape as a valuable tool to evaluate the stock structure of swordfish Xiphias gladius in the Indian Ocean.
Kélig Mahé,Djamila Ider,Andrea Massaro,Oussama Hamed,Alba Jurado-Ruzafa,Patrícia Gonçalves,Aikaterini Anastasopoulou,Angelique Jadaud,Chryssi Mytilineou,Romain Elleboode,Zohir Ramdane,Mahmoud Bacha,Rachid Amara,Hélène De Pontual,Bruno Ernande (2018). Directional bilateral asymmetry in otolith morphology may affect fish stock discrimination based on otolith shape analysis.
A Massou,P Le Bail,J Panfili,R Laë,J Baroiller,O Mikolasek,G Fontenelle,B Auperin (2004). Effects of confinement stress of variable duration on the growth and microincrement deposition in the otoliths of <i>Oreochromis niloticus</i>(Cichlidae).
S Mehanna,L Jawad,Y Ahmed,M Abu El-Regal,D Dawood (2016). Relationships between fish size and otolith measurements for<i>Chlorurus sordidus</i>(Forsskål, 1775) and<i>Hipposcarus harid</i>(Forsskål, 1775) from the Red Sea coast of Egypt.
Sahar F. Mehanna,Yassein A. A. Osman,Arafa Hassan,Magdy T. Khalil (2019). Relationships between fish and otolith dimensions of Epinephelus summana (Forsskål, 1775) and Cephalopholis argus (Schneider, 1801) from the Egyptian Red Sea coast.
F Morat,S Betoulle,M Robert,A Thailly,S Biagianti‐risbourg,R Lecomte‐finiger (2008). What can otolith examination tell us about the level of perturbations of Salmonid fish from the Kerguelen Islands?.
Fabien Morat,Yves Letourneur,David Nérini,Daniela Banaru,Ioannis Batjakas (2012). Discrimination of red mullet populations (Teleostean, Mullidae) along multi-spatial and ontogenetic scales within the Mediterranean basin on the basis of otolith shape analysis.
D Nolf (1985). Otolithi piscium: Handbook of Paleoichthyology.
A Osman,M Farrag,S Mehanna,Y Osman (2020). Use of otolithic morphometrics and ultrastructure for comparing between three goatfish species (family: Mullidae) from the northern Red Sea, Hurghada, Egypt. Iran.
D Pavlov (2016). Differentiation of three species of the genus Upeneus (Mullidae) based on otolith shape analysis.
D Pavlov,N Emel’yanova,Vo Ha,Luong Thuan (2015). Otolith morphology, age, and growth of freckled goatfish Upeneus tragula (Mullidae) in the coastal zone of Vietnam.
J Randall,D Greenfield (1999). Holocentridae: squirrelfishes (soldierfishes). FAO species identification guide for fishery purposes: the living marine resources of the Western Central Pacific: Bony fishes.
J Randall (1998). Zoogeography.
F Rohlf (2006). tpsDig 2.10.
J Rooker,D Secor,G Demetrio,A Kaufman,A Belmonte Ríos,V Ticina (2008). Evidence of trans-Atlantic movement and natal homing of bluefin tuna from stable isotopes in otoliths.
J Russ (1990). Computer-Assisted Microscopy.-The Measurement and Analysis of Image.
S Yedier,D Bostanci (2020). Aberrant otoliths in four marine fishes from the Aegean Sea, Black Sea, and Sea of Marmara (Turkey).
C Swan,M Palmer (2006). Preferential feeding by an aquatic consumer mediates nonadditive decomposition of speciose leaf litter.
V Tuset,I Lozano,J González,J Pertusa,M García-Díaz (2003). Shape indices to identify regional differences in otolith morphology of comber, <i>Serranus cabrilla</i> (L., 1758).
Victor Tuset,Antoni Lombarte,Carlos Assis (2008). Otolith atlas for the western Mediterranean, north and central eastern Atlantic.
V Tuset,A Lombarte,J González,J Pertusa,Maj. Lorente (2003). Comparative morphology of the sagittal otolith in <i>Serranus</i> spp..
Matthias Vignon (2018). Short-term stress for long-lasting otolith morphology — brief embryological stress disturbance can reorient otolith ontogenetic trajectory.
M Vignon,F Morat (2010). Environmental and genetic determinant of otolith shape revealed by a non-indigenous tropical fish.
S Yilmaz,O Yazicioglu,S Saygin,N Polat (2014). Relationships of Otolith Dimensions with Body Length of European Perch, Perca fluviatilis L., 1758 From Lake Ladik, Turkey.
No ethics committee approval was required for this article type.
Data Availability
Not applicable for this article.
How to Cite This Article
Yassein A.A. Osman. 2026. \u201cRelationship between Fish Length and otolith Shape of Sargocentron spiniferum (Forsskål, 1775 from Shalatin, Red Sea, Egypt\u201d. Global Journal of Science Frontier Research - E: Marine Science GJSFR-E Volume 23 (GJSFR Volume 23 Issue E1).
Explore published articles in an immersive Augmented Reality environment. Our platform converts research papers into interactive 3D books, allowing readers to view and interact with content using AR and VR compatible devices.
Your published article is automatically converted into a realistic 3D book. Flip through pages and read research papers in a more engaging and interactive format.
Our website is actively being updated, and changes may occur frequently. Please clear your browser cache if needed. For feedback or error reporting, please email [email protected]
Thank you for connecting with us. We will respond to you shortly.