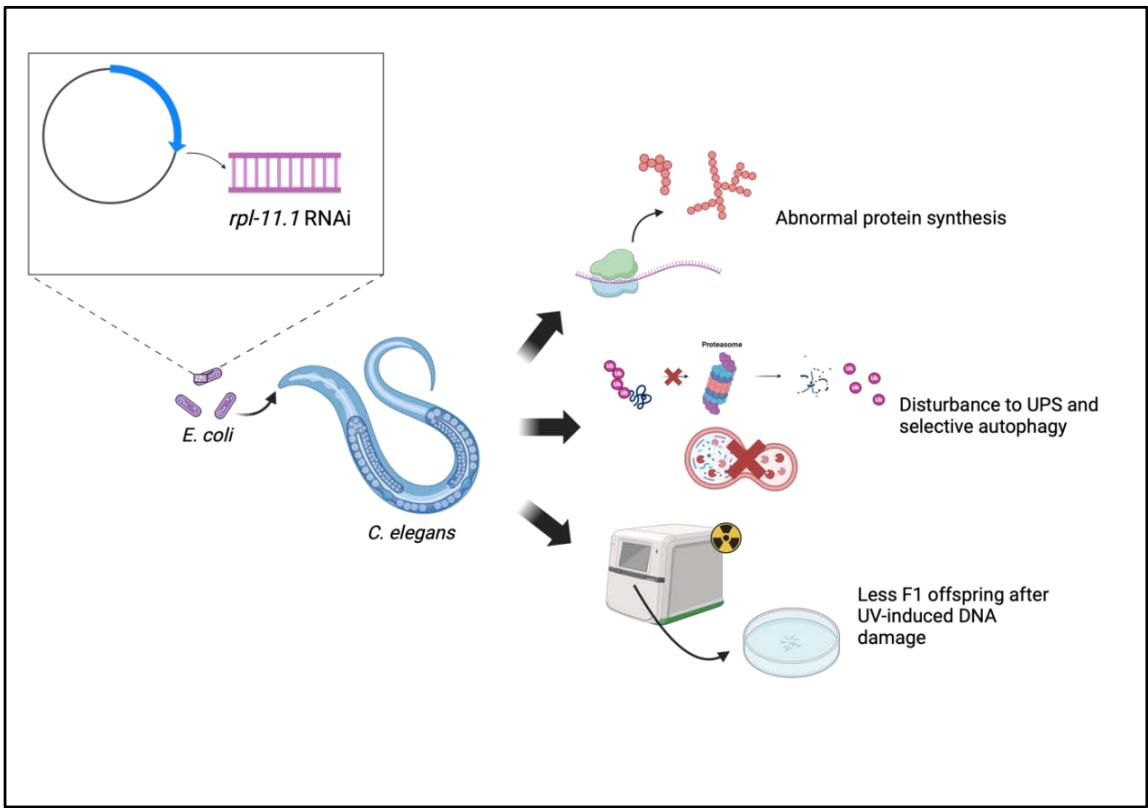
An array of human chronic diseases, such as Alzheimer’s disease, Huntington’s disease, and Parkinson’s disease, are related to defects in cellular proteostasis and the formation of protein aggregates. Using Caenorhabditis elegans PP563, a model organism developed for studying proteostasis stress, we can elucidate the biological role of specific genes and proteins involved in translation and proteostasis, advancing our understanding of relevant pathologies and therapeutics. Here, we report the knockdown of rpl-11.1 in C. elegans leads to disturbance in protein translation and proteostasis pathways, including the Ubiquitin-Proteosome System (UPS) and selective autophagy. We confirmed the importance of rpl-11.1 in ensuring correct ribosome biogenesis and translation accuracy.
## I. INTRODUCTION
Protein degradation is a major cellular process that maintains proteostasis and cellular physiology (Papaevgeniou & Chondrogianni, 2014). It primarily consists of degrading normal proteins in excessive amounts or eliminating damaged proteins, which might be a result of disturbances to normal protein synthesis in the ribosomes. A major proteostasis pathway in charge of this role is the Ubiquitin-Proteosome System (UPS), which tags damaged or misfolded proteins with ubiquitin and digests them with the 26S proteosome complex (Kipreos, 2005).
Since the genes involved in UPS are generally evolutionarily conserved, their biological roles can be investigated in simple model organisms such as Caenorhabditis elegans, a well-studied nematode for understanding fundamental biological mechanisms. A specific strain of C. elegans, PP563, has been developed to specifically investigate the UPS pathway of proteostasis. Having a GFP linked to the ubiquitin protein, C. elegans PP563 provides a direct approach to monitoring UPS activity in nematodes through fluorescence signals (McCue et al., 2015). In a genetic screen that uses RNAi to silence 35 genes at the posttranscriptional level in C. elegans PP563, we discovered a particular gene, $rpl-11.1$, whose knockdown causes a marked increase in fluorescence signal in both intensity and localization. Such phenotype led us to deduce that $rpl-11.1$ knockdown causes excessive UPS activity by increasing the number of active ubiquitin tags, which in turn disrupts proteostasis at the organismal level.
The biological role of $rpl-11.1$, a gene primarily expressed in the germline cells and pharyngeal muscle cells of C. elegans (Bgee, 2024), is still not completely elucidated. It was predicted to encode the 60S ribonucleoprotein L11-1 (RPL-11.1) in C. elegans, a homolog of ribosomal protein L11 (RPL-11) in humans (WormBase, 2022). RPL-11.1 constitutes a part of the large ribosomal subunit, which contains the peptidyl transferase center that catalyzes the formation of peptide bonds during mRNA translation (UniProt, 2024). Thereby, $rpl-11.1$ knockdown might lead to the absence of RPL-11.1 in the ribonucleoprotein complex, affecting the biogenesis of ribosomes and forming a defect in translation machinery. The incorrectly assembled ribosomes will produce misfolded proteins with altered thermodynamic stability that may be functionally damaged or prone to aggregation after exposing hydrophobic regions. An excess of misfolded proteins and protein aggregates can be the source of proteostasis disturbance observed previously: a plethora of proteins are tagged with active ubiquitin and hence emit a substantial amount of fluorescence signal.
We thereby hypothesize that knocking down $rpl-11.1$ via RNAi in C. elegans results in structurally incomplete ribosomes, which might produce misfolded proteins, recruit excessive ubiquitin for tagging and clearing through the UPS pathway, and ultimately overwhelm the proteostasis. Answering this hypothesis would reveal the biological role of $rpl-11.1$ more clearly and possibly shed light on its relationship with translation efficiency and proteostasis regulation. To verify the hypothesis, two aims have been formulated: first, to determine if $rpl-11.1$ has a role in ensuring correct protein synthesis. This will be achieved by conducting protein assays such as SDS-PAGE to compare the pattern of protein expression in $rpl-11.1$ knockdown and that in C. elegans PP563 on an empty vector (L4440). We will further examine whether the other pathway of proteostasis that also makes use of ubiquitin tagging, selective autophagy, is involved in the clearance of $rpl-11.1$ knockdown-induced protein aggregates and misfolded proteins as well. This would require knocking down $rpl-11.1$ in C. elegans MAH215, which is built for studying autophagy (Chang et al., 2017), and conducting a double-gene RNAi knockdown in C. elegans PP563 for comparison with $rpl-11.1$ single knockdown. Another minor aim of this study would be to determine the role of $rpl-11.1$ in protecting germline cells. The human orthologue of C. elegans RPL-11.1 is involved in p53 pathway regulation through RPL11-MDM2 antagonism and acts as a tumor suppressor (Chène, 2003). Similarly, C. elegans also has a p53-like pathway (cep-1) that regulates DNA damage-induced apoptosis in germline cells (Derry, 2001). We hypothesize that knocking down $rpl-11.1$ will inhibit cep-1 stabilization and hence germline apoptosis after UV-induced DNA damage, allowing increased germline cell proliferation and more egg deposition. This will be investigated through a F1 progeny count following parent generation exposure to UV.
As mentioned, the functionality and mechanism of $rpl-11.1$ in C. elegans is not fully clarified in literature. Some of the earliest genome-wide screens indicate that $rpl-11.1$ knockdown resulted in phenotypes such as life-span extension(Hsin & Kenyon, 1999) and gigantism(Patel et al., 2002). More recent studies have focused on the germline proliferation aspect of the gene as well as its evolutionary history in the C. elegans genome(Maciejowski et al., 2005). A more interesting study, without any experimental evidence, suggested the potential involvement of the gene in mitochondrial activity.(Sun et al., 2019). These various studies seemingly covered the role of $rpl-11.1$ in C. elegans from a broad perspective, but none of them point out the specific mechanism by which this gene works in terms of its fundamental role: encoding a ribosomal protein. Additionally, none of the prior research exclusively focused on studying $rpl-11.1$. Here, we solely investigated $rpl-11.1$ knockdown from the perspective of translation and proteostasis disturbance and germline proliferation. Elucidating the biological role of $rpl-11.1$ in nematodes is of great importance as it can give insight into similar mechanisms in the translation machinery and UPS pathway of humans. A substantial homogeneity exists between humans and C. elegans(Lai, 2000), so confirming the gene's role inthe nematode can potentially pave the way to developing therapeutic strategies for diseases related to protein aggregation and irregular proteostasis, such as Alzheimer's disease,
Huntington's disease, Parkinson's disease, and prion disorders(Papaevgeniou & Chondrogianni, 2014).
## II. METHODS
Culture of C. elegans. C. elegans PP563 and C. elegans MAH215 were used in this study and obtained from the Caenorhabditis Genetics Center (University of Minnesota, St. Pau, MN, USA). They were maintained at $25^{\circ}\mathrm{C}$ under standard conditions on nematode growth media (NGM; $2\%$ (w/v) agar, $0.3\%$ (w/v) NaCl, $0.25\%$ (w/v) peptone, $1\mathrm{mM}$ $\mathrm{CaCl}_2$, $5\mu \mathrm{g}$ $\mathrm{ml}^{-1}$ cholesterol, $25\mathrm{mM}$ $\mathrm{KH}_2\mathrm{PO}_4$, $1\mathrm{mM}$ $\mathrm{MgSO}_4$ ) agar plates(Sun et al., 2019). Nematodes were fed on E. coli OP50.
PP563 RNAi screen. The following screen protocol was followed:
Day 1. $100 \mathrm{~uL}$ of cultures of the Htt115 strain of $E.$ coli containing the L4440 plasmid alone or containing an RNAi sequence targeting a specific gene were seeded onto $\mathrm{NGM}+$ carbenicillin p6 plates and allowed to dry overnight. Each plate has one targeting RNAi. (Refer to these plates as RNAi p6s.)
Day 2. All RNAi p6s had $50~\mu \mathrm{L}$ of IPTG added to enhance expression of the RNAi. This was allowed to dry ( $\sim$ 1 hour). Gravid worms were bleached to obtain a synchronized population of eggs. Approximately 50 eggs were plated onto the RNAi p6s and were allowed to develop to adulthood ( $\sim$ 3 days) at $20^{\circ} \mathrm{C}$.
Day 5. About 10 worms were randomly selected from each RNAi p6, immobilized with sodium azide, and aligned for imaging. Micrographs were imaged at 150x magnification and 20ms exposure with LED at 16 and white-balanced. For the GFP signal, C. elegans PP563 was imaged at 800 ms exposure, and C. elegans MAH215 was imaged at 500 ms exposure. For the mCherry signal, C. elegans MAH215 was imaged at 500 ms exposure.
We conducted a preliminary screen for C. elegans PP563 with 36 different RNAi knockdowns. Of all the knockdowns that showed some degree of change in fluorescence signal in comparison to the L4440 genetic control, the rpl-11.1 knockdown displayed the most drastic increase in both signal intensity and localization. We therefore decided to follow up on our investigation on the RPL-11.1 knockdown exclusively.
The knockdown efficiency of $rpl-11.1$ RNAi was confirmed using RT qPCR, where the mRNA level of the housekeeping gene ama was used as the internal reference. Primers for the target genes were designed by primer-BLAST (NCBI, 2019) and synthesized by a commercial company. The RT qPCR result showed that $rpl-11.1$ RNAi achieved a near-complete (0.0025 fold change) knockdown of $rpl-11.1$ in C. elegans PP563. It should be noted that the same gene knockdown in C. elegans MAH215 was not verified by RT qPCR.
Nematode population maintenance: Nematodes were synchronized by hypochlorite bleaching (2% sodium hypochlorite and 0.5 mol/L NaOH) according to standard protocols (Stiernagle, 2006). Approximately 1,000 extracted worm eggs were cultured on one NGM plate for protein isolation and RNA isolation (for RT qPCR cDNA synthesis).
Protein isolation: Proteins from C. elegans PP563 rpl11.1 knockdown and L4440 are isolated by washing the worms off the plate following standard protocols (Stiernagle, 2006), lysing through RIPA (50mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 1% or 5% SDS, tablet of protease inhibitor), centrifugation and incubation on ice. Proteins treated with 1% SDS are generally small-sized and soluble, while the 5% SDS aims to separate out proteins that are large-sized or aggregated.
SDS-PAGE: RIPA-treated proteins (including $1\%$ SDS-treated and $5\%$ SDS-treated) were loaded onto protein gel to run SDS-PAGE at 170V for the first 10 minutes and then at 200V for 1 hour. The gel was destained and visualized in imaging system.
Rpl-11.1 RNAi knockdown in C. elegans MAH215: C. elegans MAH215 were fed with rpl-11.1 RNAi-expressing bacteria during development. Procedure similar to PP563 RNAi screen was followed. Micrographs were imaged at 31.6 ms exposure and 2x gain. GFP and mCherry fluorescence signal are later overlayed to create merged fluorescence micrographs.
Double-gene RNAi knockdown: Two genes (rpn-6 and lgg-1, rpl-11.1 and lgg-1, rpn-6 and rpl-11.1) were knocked down via RNAi at once in C. elegans PP563 to evaluate the involvement of UPS and autophagy pathways of proteostasis in translation-defect cells' protein degradation. Bacteria expressing both types of RNAi are used as food source for worms and the procedure is similar to PP563 RNAi screen. Micrographs were imaged at 31.6 ms exposure and 2x gain.
UV radiation exposure: Two plates of $rpl-11.1$ knockdown C. elegans PP563 and two plates of L4440 C. elegans PP563 were placed under a UV source (imaging system), with one plate exposed to UV for 30 sec and the other for 120 sec. Then, 5 adult worms from each plate is transferred to new plates seeded with E. coli OP50 and cultured for 3 days at $25^{\circ}\mathrm{C}$ till the F1 progeny reaches adulthood. The number of worms on each plate was then counted manually. After worm (population) count, 10 worms from each group were transferred to new plate, immobilized and imaged under fluorescent microscope.
## III. RESULTS
a) $5\%$ SDS-PAGE Reveals Potential Protein Aggregation In Figure 1, it is observed that under $1\%$ SDS treatment, which separates out smaller proteins, there is no clear difference between the bands of $rpl-11.1$ knockdown and the control (L4440). Nonetheless, under $5\%$ SDS treatment that targets larger proteins, $rpl-11.1$ knockdown exhibits markedly more bands and higher intensity in comparison with the control. These extra bands might be protein aggregates accumulated in the cells.
### b) Rpl-11.1 Knockdown in MAH215 Induces More Merged Fluorescence Signal
In Figure 2, merged micrographs generally reveal more stand-alone m Cherry fluorescence in the control (L4440) and more merged fluorescence (mCherry and GFP) in $rpl-11.1$ knockdown. One exception is that there are two nematodes in $rpl-11.1$ knockdown that exhibit GFP fluorescence unaccompanied by any mCherry fluorescence. These two nematodes were believed to be dead by the time of imaging and were displaying auto-fluorescence because the GFP signal must be accompanied by the mCherry signal in C. elegans MAH215. Other than the two abnormalities, $rpl-11.1$ knockdown displays mostly merged fluorescence.
It should be noted that in an attempt to verify if the autophagy pathway of proteostasis is involved in misfolded protein clearance, a double-gene knockdown experiment was also performed (refer to Methods). However, no valid results were obtained because there is minimal difference between the control and experiment groups (Figure 3).
### c) Rpl-11.1 Knockdown Produces Less F1 Progeny after UV Exposure
Figure 4A reveals that there is no obvious difference between UV-treated $rpl-11.1$ knockdown and the control (L4440) in terms of F1 progeny phenotype (fluorescence, size, mobility, etc.). However, when a worm count was conducted, it turned out that in both the 30s and 120s UV exposure groups, there are more nematodes surviving in the control group, with a 6.4-fold increase in the 30s exposure and a 2.5-fold increase in the 120s exposure. The knockdown of $rpl-11.1$ inhibits nematode proliferation after UV exposure.
## IV. DISCUSSION
### a) Protein Aggregation as a Result of Error in Translation Machinery
Our SDS-PAGE experiment substantiated that there are protein aggregates formed in C. elegans PP563 cells after rpl-11.1 knockdown, as indicated by extra bands of proteins that are not seen in the control (Figure 1). Moreover, although the expression of smaller proteins appears to be similar in both the control and the rpl-11.1 knockdown, we observed an overexpression of larger proteins (5% SDS-treated) in the rpl-11.1 knockdown, suggesting a difference in protein expression pattern. This aligns with our hypothesis that abnormal protein synthesis will arise due to the absence of RPL-11.1 ribonucleoprotein in ribosome biogenesis. The abnormality can take the form of producing misfolded polypeptides that have altered thermodynamic stability, which are prone to exposing their hydrophobic regions and hence become aggregated (Berrill et al., 2011). Alternatively, the abnormal protein synthesis might also simply result in an overexpression of a group of large, insoluble proteins. Both outcomes will cause severe disruption to the UPS because excessive amounts of ubiquitin will be tagged to abnormal proteins, overwhelming the proteostasis pathway.
It is confirmed that knocking down $rpl-11.1$ makes a difference in cellular protein synthesis in C. elegans. Next, we will attempt to identify which specific proteins are affected (overexpressed, misfolded, or become prone to aggregation) by such errors in translation machinery. Our preliminary RNAi screen indicated that most proteostasis defects occur in C. elegans intestinal cells after knocking down $rpl-11.1$. We will select a set of proteins whose expressions may be disturbed by the knockdown and use immunoblotting to confirm their identity. In addition, we will evaluate the degree of impact on ribosome biogenesis and translation efficiency caused by knocking down $rpl-11.1$ through RT qPCR on rRNAs and polysome profiling, respectively.
### b) Autophagy Pathway is Disturbed by Rpl-11.1 Knockdown Alongside UPS
Alongside UPS, the selective autophagy pathway of proteostasis also makes use of ubiquitin tagging to achieve protein degradation (Kocaturk & Gozuacik, 2018). We indeed observed a disturbance to the autophagy pathway in the C. elegans MAH215 rpl-11.1 knockdown (Figure 2). The abundant mCherry signal in the control suggests that most GFP tagged to the autophagosomes has been quenched after fusing with the lysosome in the autophagy pathway. Conversely, in rpl-11.1 knockdown, an increase in merged fluorescence signal indicates that autophagosomes are not efficiently undergoing lysosomal fusion, suggesting a delay in protein degradation and a buildup of protein waste. This is likely attributable to the protein aggregates and other large misfolded proteins generated by the translation error. Therefore, the hypothesis that selective autophagy is also involved in this knockdown-induced proteostasis disturbance is supported. Interestingly, autophagy differs from the UPS in that it primarily degrades long-lived proteins, insoluble protein aggregates, and organelles, whereas the UPS pathway targets short-lived proteins and soluble misfolded proteins (Kocaturk & Gozuacik, 2018). This aligns with our findings from the protein assay, which show that knocking down rpl-11.1 produces protein aggregates and other insoluble large proteins, necessitating the involvement of selective autophagy in maintaining proteostasis.
Our hypothesis could be better supported with results from the double-gene knockdown experiment. If simultaneously knocking down $rpl-11.1$ and $lgg-1$, a key gene involved in autophagy (Romane Leboutet et al., 2023), produces a result that is no different from knocking down $rpl-11.1$ alone, we could conclude that the autophagy pathway of proteostasis is already disturbed by knocking down $rpl-11.1$ in addition to the UPS. However, unfortunately, the experiment did not yield valid data to draw any conclusions. We suspect that the RNAi knockdown efficiency might be problematic, resulting in an incomplete silencing of the targeted gene. To ensure validity in re-performing the double-gene knockdown experiment, we will include a confirmation of RNAi knockdown efficiency using RT qPCR.
### c) Rpl-11.1 Might Be Integral to Germline Proliferation
We previously predicted that knocking down $rpl-11.1$ will lead to more F1 offspring as germline apoptosis is inhibited by the deactivation of cep-1, which is under RPL-11.1 regulation(Schumacher et al., 2001). However, the experiment result indicates that $rpl-11.1$ knockdown produces much less F1 progeny in comparison with the control after UV exposure for either 30s or 120s. The original hypothesis is hence rejected. We need to reconsider the role of $rpl-11.1$ in maintaining germline proliferation.
According to Chang et al. (2017), cep-1, while activating DNA damage-induced germline apoptosis, is also required for meiotic chromosome segregation in the germline. Hence, it is reasonable that knocking down $rpl-11.1$, the stabilizer of cep-1, leads to less progeny after UV-induced DNA damage in the parent generation. Also, the mechanism by which cep-1 is stabilized by RPL-11.1 in C. elegans might be different from how p53 (the human homolog of cep-1) is stabilized by RPL-11 (the human homolog of RPL-11.1) in humans, requiring us to figure out the specific mechanism of cep-1 activation before making predictions. It can be concluded that $rpl-11.1$ might be integral to germline proliferation in C. elegans in the face of radiation stress, based on the UV exposure experiment result.
In further investigation, we attempt to monitor the process of meiotic chromosome segregation in the germline of $rpl-11.1$ knockdown C. elegans using live microscopy imaging. We will observe the gonads of the nematodes under fluorescent microscopes after their chromosomes are stained with fluorescent markers. This will allow us to evaluate our new hypothesis.
In conclusion, our original hypothesis was partially supported. The role of $rpl-11.1$ in ensuring correct protein synthesis in C. elegans is confirmed. We believe the selective autophagy pathway of proteostasis is involved in the clearance of misfolded proteins alongside UPS, but evidence from the double-gene RNAi knockdown experiment is lacking. On the other hand, the role of $rpl-11.1$ in protecting germline cells requires re-consideration and further investigation. It appears that the gene may be a necessity for meiotic chromosome segregation, as it plays an important role in nematode reproduction after radiation-induced DNA damage.
Due to the substantial homogeneity between humans and C. elegans (Lai, 2000), it is of utter importance to continue elucidating the biological role of rpl-11.1 and its protein product in translation and cellular proteostasis, which relates closely to an array of human diseases involving proteomic defects. We would like to further evaluate the disturbance caused by RPL-11.1 dysfunction to translation efficiency by conducting polysome profiling and rRNA qPCR. The immunoblotting of specific proteins involved in the disturbance is equally crucial for developing therapeutic strategies toward relevant proteomic diseases.
#### ACKNOWLEDGEMENT
We would like to thank Dr. Mills for her invaluable guidance in research design and support in experimental work throughout this research. We are also grateful to Dennis Bonal and Callie Millette for their assistance in our lab work and for correcting any issues in our research with patience. We express our gratitude to the MAH215 group for providing us with the MAH215 strain for conducting our investigation and to all other groups for their insightful advice given to our research design. Last but not least, special thanks to the MDL staff for providing us all the necessary equipment and materials for our experiments, without which we could not have completed this research.
Generating HTML Viewer...
References
20 Cites in Article
Alex Berrill,Jamie Biddlecombe,Daniel Bracewell (2011). Product Quality During Manufacture and Supply.
Jessica Chang,Caroline Kumsta,Andrew Hellman,Linnea Adams,Malene Hansen (2017). Spatiotemporal regulation of autophagy during Caenorhabditis elegans aging.
P Chène (2003). Inhibiting the p53-MDM2 interaction: an important target for cancer therapy.
W Derry,Aaron Putzke,Joel Rothman (2001). <i>Caenorhabditis elegans</i> p53: Role in Apoptosis, Meiosis, and Stress Resistance.
Honor Hsin,Cynthia Kenyon (1999). Signals from the reproductive system regulate the lifespan of C. elegans.
Edward Kipreos (2005). Ubiquitin-mediated pathways in C. elegans.
N Kocaturk,D Gozuacik (2018). Crosstalk Between Mammalian Autophagy and the Ubiquitin-Proteasome System.
Chun-Hung Lai,Chang-Yuan Chou,Lan-Yang Ch'ang,Chung-Shyan Liu,Wen-Chang Lin (2000). Identification of Novel Human Genes Evolutionarily Conserved in<i>Caenorhabditis elegans</i>by Comparative Proteomics.
John Maciejowski,James Ahn,Patricia Cipriani,Darrell Killian,Aisha Chaudhary,Ji Lee,Roumen Voutev,Robert Johnsen,David Baillie,Kristin Gunsalus,David Fitch,E Hubbard (2005). Autosomal Genes of Autosomal/X-Linked Duplicated Gene Pairs and Germ-Line Proliferation in Caenorhabditis elegans.
H Mccue,X Chen,J Barclay,A Morgan,R Burgoyne (2015). Expression profile of a Caenorhabditis elegans model of adult neuronal ceroid lipofuscinosis reveals down regulation of ubiquitin E3 ligase components.
(2019). Table 1: Reference gene qPCR primers designed using NCBI Primer BLAST..
N Papaevgeniou,N Chondrogianni (2014). The ubiquitin proteasome system in Caenorhabditis elegans and its regulation.
Mavji Patel,Christopher Knight,Constantina Karageorgi,Armand Leroi (2002). Evolution of germ-line signals that regulate growth and aging in nematodes.
Romane Leboutet,Céline Largeau,Leonie Müller,Magali Prigent,Grégoire Quinet,Manuel Rodriguez,Marie-Hélène Cuif,Thorsten Hoppe,Emmanuel Culetto,Christophe Lefebvre,Renaud Legouis (2023). LGG-1/GABARAP lipidation is not required for autophagy and development in Caenorhabditis elegans.
Björn Schumacher,Kay Hofmann,Simon Boulton,Anton Gartner (2001). The C. elegans homolog of the p53 tumor suppressor is required for DNA damage-induced apoptosis.
T Stiernagle (2006). Maintenance of C. elegans. WormBook: The online review of C. elegans biology.
Yongyan Sun,Xiaomei Huang,Yahong Wang,Zhenhua Shi,Yanyan Liao,Peng Cai (2019). Lipidomic alteration and stress-defense mechanism of soil nematode Caenorhabditis elegans in response to extremely low-frequency electromagnetic field exposure.
(2024). UniProt: Programmatic access to UniProtKB: webinar.
Zishuo Sam Li,Jocelyne Mills,Dennis Bonal,Callie Millette (2024). rpl-11.1 Knock-Down Disturbs Translation Machinery and Proteostasis in Caenorhabditis elegans.
Explore published articles in an immersive Augmented Reality environment. Our platform converts research papers into interactive 3D books, allowing readers to view and interact with content using AR and VR compatible devices.
Your published article is automatically converted into a realistic 3D book. Flip through pages and read research papers in a more engaging and interactive format.
An array of human chronic diseases, such as Alzheimer’s disease, Huntington’s disease, and Parkinson’s disease, are related to defects in cellular proteostasis and the formation of protein aggregates. Using Caenorhabditis elegans PP563, a model organism developed for studying proteostasis stress, we can elucidate the biological role of specific genes and proteins involved in translation and proteostasis, advancing our understanding of relevant pathologies and therapeutics. Here, we report the knockdown of rpl-11.1 in C. elegans leads to disturbance in protein translation and proteostasis pathways, including the Ubiquitin-Proteosome System (UPS) and selective autophagy. We confirmed the importance of rpl-11.1 in ensuring correct ribosome biogenesis and translation accuracy.
Our website is actively being updated, and changes may occur frequently. Please clear your browser cache if needed. For feedback or error reporting, please email [email protected]
Thank you for connecting with us. We will respond to you shortly.