



I. INTRODUCTION
Mycorrhizae are symbioses between plant roots and certain soil fungi. The term mycorrhiza describes the group made up of the absorption structures of plants and the fungal symbionts that colonize them in a distinctive way (Sánchez de Prager, 2007; Perez et al., 2010), generating a structural and metabolic integration typical of a symbiosis. The absorption of nutrients is affected by these associations that also influence plant health, productivity, and the adaptation of plants to environmental conditions (Sánchez de Prager, 2007).
The plant provides the fungus with carbohydrates, proteins, and lipids, necessary for its development, and in turn, the fungus allows the plant to better capture water and nutrients from the soil, both macronutrients and micronutrients, especially those few mobile, such as phosphorus (Sánchez de Prager, 2007).
Endo mycorrhizae and ectomycorrhizae are two types of mycorrhizae that occur naturally (Sieverding, 1983; García et al., 2000; Biaus, 2017). Endomycorrhizae are more frequent in nature and are characterized by the colonization of cortical cells by a mycosymbiont, which lives between them and within them, inter and intracellularly (Montenegro Gómez et al., 2017). Among the different types of endomycorrhizal, the most widespread within plant species are vesicular-arbuscular mycorrhizae (VAM) or arbuscular mycorrhizae (AM) and some more specific forms such as orchidoids and ericoid (Sánchez de Prager, 2007; Montenegro Gómez et al., 2017).
Arbuscular mycorrhizal fungi (AMF) and vesicular-arbuscular fungi (HMVA) do not develop in pure culture media because they are not capable of growing in the absence of a host plant (Aguilar-Ulloa et al., 2016), so they are considered obligate symbionts, this biological condition that represents an obstacle to their massive propagation. These fungi are propagated using trap plants, which after a period of growth, the roots and soil are collected for use as inoculant of endomycorrhizal fungi (De la Rosa-Meraet al., 2012). In recent decades, the study of these organisms has become important for apply them to the ground, as biofertilizers to increase the productivity in crops (Covacevich and Echeverria, 2010; Reyes Tena et al., 2015; Aguilar-Ulloa et al., 2016; Ordoñez-Castaneda et al., 2021) and in phytoremediation, programs to correct contaminated soils (Pérez et al., 2021; Colombo et al., 2020; Quiroz-Mojica et al., 2021). The colonization of roots by endomycorrhizal fungi is the most used parameter as a quick indicator of the presence of mycorrhizal symbiosis (Covacevich and Echeverria, 2010).
Several plant species have been investigated as trap crops to produce mycorrhizae, to find plants that can be colonized and allow the rapid growth of mycorrhizae (Aguilar-Ulloa et al., 2016). The appropriate trap crop must be a fast-growing plant that adapts to the environmental conditions where it will grow, must be easily colonized by the mycorrhizal fungus, and produce many roots in a relatively short time (45-60 days) (Siqueira Martins et al., 2017).
The association established by AMF is not specific, which allows the same fungus to colonizes different plant species to generate symbiosis (Aguilar-Ulloa et al., 2016). Also, there is a preference that certain AMF colonize and spread better in certain plant species (Covacevich and Echeverria, 2010). Furthermore, using the same fungal species is probably not optimal for all crops. For this reason, it is convenient to evaluate the MVA in each trap crop species.
Because the interactions between different VMAs with soil microorganisms are complex, it is necessary to determine their behavior in the field under the growth conditions of each crop. The use of native MVA is recommended due to its adaptation to prevailing conditions, avoiding ecological risks associated with introducing exotic species (Sánchez de Prager, 2007).
The objective was to determine the existence of mycorrhizal associations between autochthonous soil fungi and forage crops planted in the Central Valley of Catamarca and to evaluate the biological aspects of the interactions found.
II. MATERIALS AND METHODS
Colonization of mycorrhizae in plant roots of six commonly cultivated forage species in the region was studied. The sowings were carried out in the locality of Miraflores, Department of Capayán, in the Central Valley of Catamarca, in plots of native forest or with agricultural history without application of mycorrhizal fungi. The species evaluated were: clover (Melilotus officinalis L.), oats (Avena sativa L.), barley (Hordeum vulgare L.), rye (Secale cereale L.), Gatton panic (Panicum maximum Jacq.), and buffel grass (Cenchrus ciliaris L.).
In the laboratory, the roots of each of the collected plants were extracted and washed with running water. The thinnest were selected, those that were clarified and stained following the methodology of Phillips and Hayman (1970) to determine fungal colonization and detection of mycorrhizal structures. Staining was performed with Gueguén's triple dye solution, allowing fungi to stain proteins blue simultaneously, starch violet, fats red (Sarasola and Rocca, 1975), and glycogen in mahogany (Verna and Herrero, 1952).
In each specimen, the percentage of fungal colonization by MVA and the percentages of the content of arbuscules (A) and vesicles (V) were quantified by the line intersection method of Giovannetti and Mosse (1980), and the frequency of appearance of A and V (Covacevich et al., 2001). For each specimen, 15 segments of the colored roots were taken, and distributed randomly on a grid slide. By means of microscopic observation (10x and 40x), the presence absence of mycorrhizal structures (A and V) was recorded in the horizontal and vertical intersections between roots and grid lines. Three repetitions of each species were performed, quantifying at least 100 intersections per preparation to later calculate the frequency of mycorrhizal infection, according to Giovannetti and Mosse (1980).
MGiovannetti\,\%\,=\,\text{No.\,SI}\,\times\,100\,\/\,\text{No.\,SO}Where:
SI: number of infected segments (hyphae + arbuscules + vesicles) and
SO: numberof total segments observed (hyphae + arbuscules + vesicles + no infection).
Also, the percentage of hyphae of dark septateendophytes (ESO) and the percentage of microsclerotia were recorded.
III. RESULTS AND DISCUSSION
In the six analyzed forage species, typical structures of endomycorrhizal fungi of the MVA type (hyphae, arbuscules, and vesicles), and of ESO fungi (with septate, melanized hyphae, and numerous microsclerotia) were observed.
In all species there were continuous thin and thick hyphae, with intracellular and intercellular growth and some of them with lipids in a rosary inside.
In clover, arbuscular distributed throughout the bark were observed, involved in the bidirectional transfer of nutrients (Smith and Read, 1997). Numerous vesicles of diverse morphology (spherical, oval, tapered) were also observed in this legume, so it is inferred that the roots are colonized by various species or genera of native HMVA. Vesicles with light blue (saccule) and red (single or multiple globules) colorations were observed. These structures are related to the carbon storage of in the form of lipids and fatty acids. For this reason, vesicles are defined as reserve organs of the fungal symbiont (Sieverding, 1983).
A high level of fungal colonization was determined in the six forage plants, obtaining the highest values of mycorrhizal colonization (MC) in clover and barley roots (Table 1). The highest frequency of appearance of arbuscules (FAA) was observed in clover, while the lowest number of arbuscules was determined in oats. The highest frequency of vesicle appearance (FAV) was also observed in clover, with average values of (Table 1). These results coincide with the statements of Covacevich and Echeverria (2010) that indicate that there is a preference for certain AMF to better colonize certain plant species. The more significant colonization in clover is explained by the fact that Rhizobios-arbuscular mycorrhizae (AM) associations occur in legumes that act synergistically in infection, mineral nutrition, and plant growth (Fitter & Garbaye, 1994; Barea, 1997). The main effect of AM in enhancing rhizobia, activity is through a generalized stimulation of plant nutrition, some more localized
effects may occur at the root (Melo de Miranda et al., 2008; Spagnolletti et al., 2021).
| Species | Mycorrhizal Fungi | Fungi ESO FAM4 (%) | ||
| MC1 (%) | FAA2 (%) | FAV3 (%) | ||
| Clover | 89 | 47 | 20 | 15 |
| Barley | 89 | 17 | 15 | 15 |
| Rye | 83 | 27,7 | 16 | 13 |
| Buffalograss | 76,5 | 20 | 8 | 2 |
| gatton panic | 75,8 | 32 | 9,8 | 1,7 |
| Oatmeal | 75 | 10 | 12 | 4 |
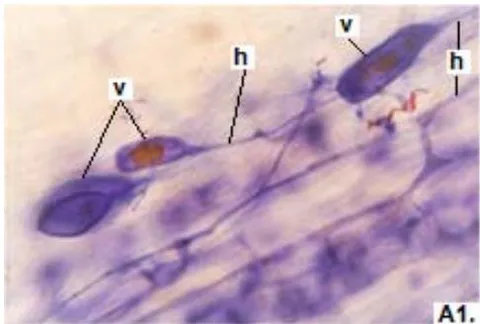
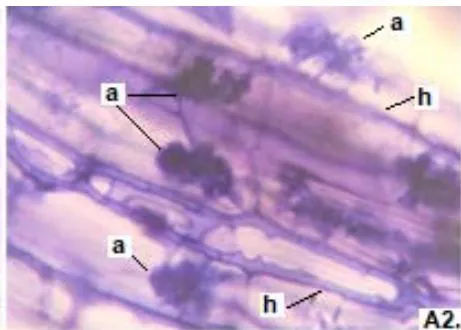
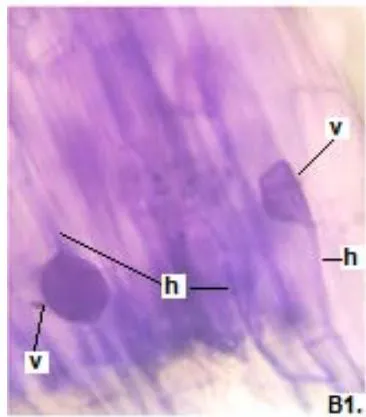
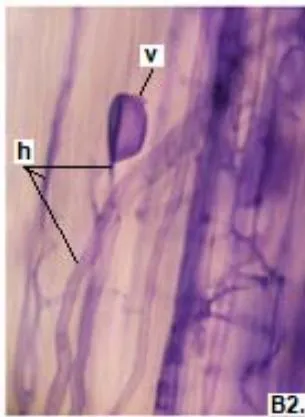
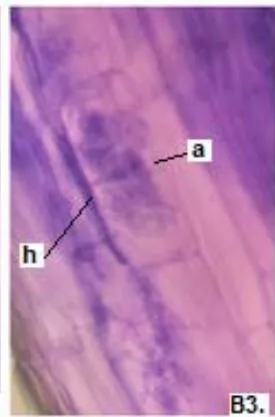
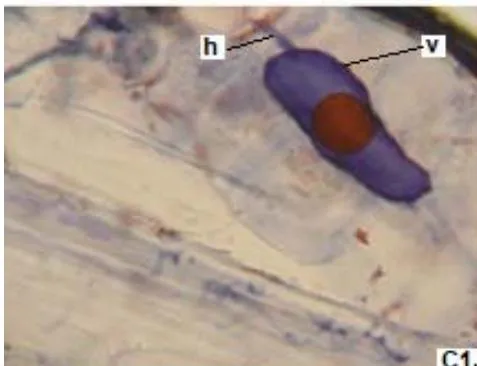
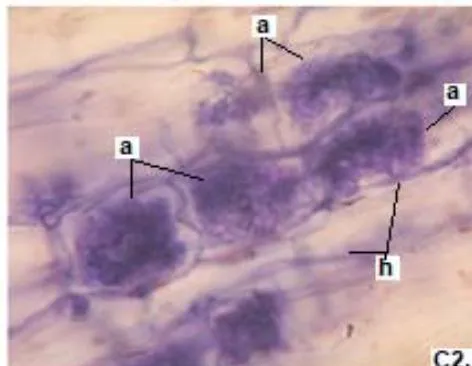
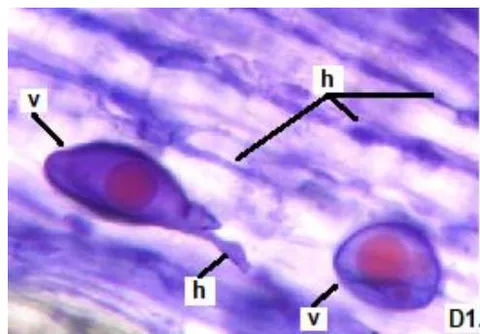
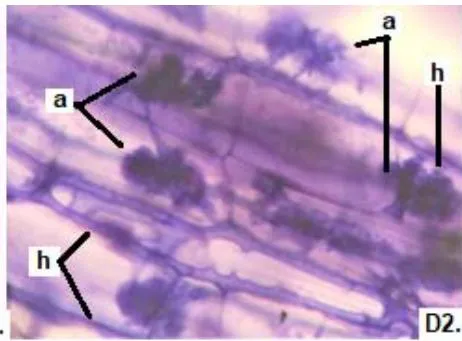
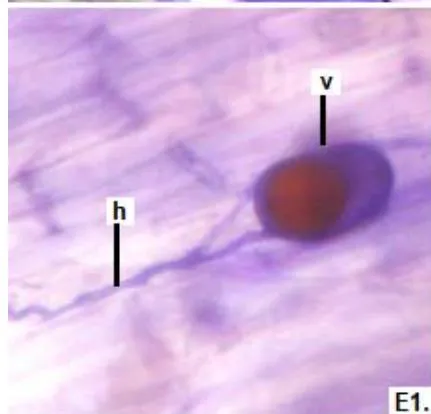
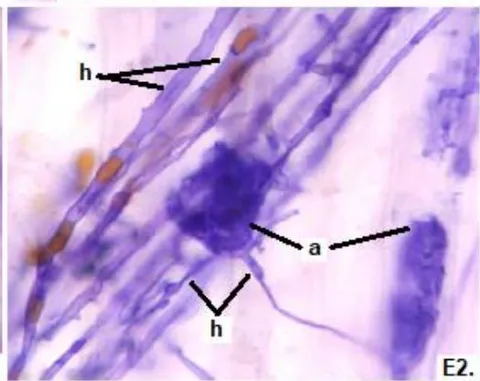
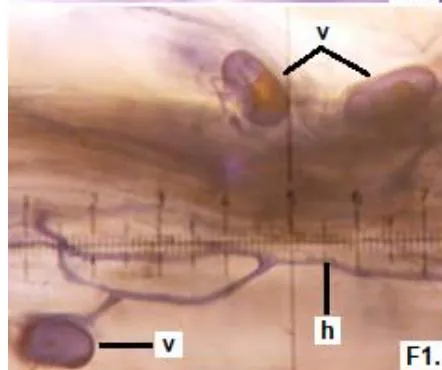
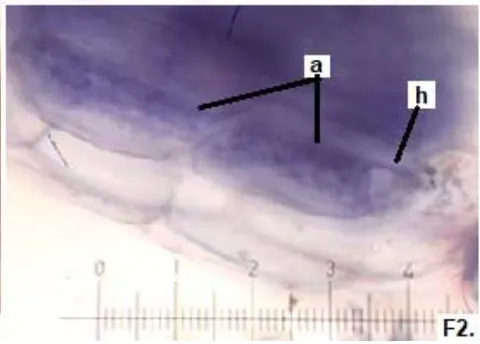


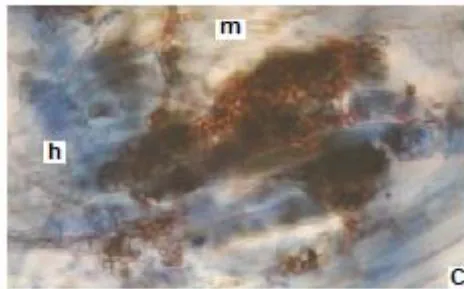
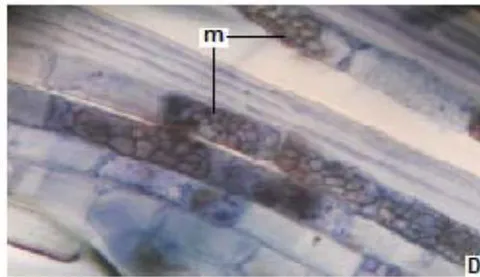


In all the plants studied, their roots were simultaneously colonized by both endophytes, MVA and ESO hyphae, also detecting the presence of vesicles, arbuscules typical of VA mycorrhizae, and ESO microsclerotia. This was also observed in other plants including ferns, mono, and dicots that are simultaneously colonized by MVA and ESO. (Urcelay et al., 2005; Lugo et al., 2011; Lizárraga et al., 2015; Di Barbaro et al., 2017).
Therefore, MVA and other fungal structures developed in all the forage species studied. This coincidence could be due to the fact that all these crops were carried out in contiguous lots with identical soil characteristics and environmental conditions, and because the fungal colonization developed from the native microflora, with the ability to associate and generate MVA, which is consistent with what expressed by Aguilar-Ulloa et al. (2016) where they explain that the same fungus can colonize different plant species to generate symbiosis.
These mycorrhizal-forming fungi can be considered as potential constituents of biofertilizers. Diaz Franco et al. (2019) achieved the reduction of inorganic fertilization through the inoculation of FAM in the sorghum crop. The higher yield of sorghum obtained allows considering the inclusion of FAM as a viable practice that guarantees greater profitability, as well as the agroecological conservation of production systems.
IV. CONCLUSION
The association of mycorrhizae in six forage species cultivated in the Central Valley of Catamarca and the co-occurrence of vesicular-arbuscular mycorrhizae and native dark septate endophytes is described.
High levels of mycorrhizal colonization were obtained in all the evaluated forage species. The suitability of Melilotus officinalis as a trap species is highlighted as it is easily colonized by mycorrhizal fungi and generates rapid mycorrhizal growth.



